Complete clinical study: Zuzzi-Krebitz AM, Buchta K, Bergmann M, Krentz D, Zwicklbauer K, Dorsch R, Wess G, Fischer A, Matiasek K, Hönl A, Fiedler S, Kolberg L, Hofmann-Lehmann R, Meli ML, Spiri AM, Helfer-Hungerbuehler AK, Felten S, Zablotski Y, Alberer M, Both UV, Hartmann K. Short Treatment of 42 Days with Oral GS-441524 Results in Equal Efficacy as the Recommended 84-Day Treatment in Cats Suffering from Feline Infectious Peritonitis with Effusion-A Prospective Randomized Controlled Study.Viruses. 2024 Jul 16;16(7):1144. doi: 10.3390/v16071144. PMID: 39066306; PMCID: PMC11281457.
The discovery of GS-441524 as an effective antiviral drug for cats with feline infectious peritonitis (FIP) has enabled feline patients to survive this once incurable, fatal disease. In the UK and Australia, GS-411524 is already legally available, while in the US the drug has only recently been available through selected compounding pharmacies. An 84-day treatment cycle has been shown to be successful in various clinical studies and has become an unofficial standard protocol. From a practical point of view, the daily administration of the drug for 12 weeks, as well as the cost of such treatment, can make it difficult or even impossible for cat owners to complete the entire prescribed treatment.
The aim of the researchers in Germany and Switzerland was to evaluate whether a 42-day treatment with GS-441524 is as effective as the currently recommended 84-day protocol. In a prospective randomized controlled treatment study, 40 cats were randomized to receive 15 mg/kg GS-441524 orally once daily for 42 or 84 days. Patients were diagnosed with FIP based on either FCoV RNA detected by RT-qPCR or RT-PCR in effusion in at least one body cavity with altered laboratory parameters typical of FIP. In addition to the diagnosis of FIP, other inclusion criteria included the presence of abdominal and/or pleural effusion, negative FeLV and FIV status, a body weight of at least 2 kg, and the absence of other serious diseases. The age of the cats ranged from 5.1 to 116.3 months, with 17 of the 40 cats being less than 1 year old. Breed distribution was as follows: 40 % Domestic Shorthairs (DSH), 20 % British Shorthairs (BSH) and 40 % other breeds. At the start of the study, 63 % cats had abdominal effusion, 12 % pleural effusion, and 25 % effusion in both cavities.
Each patient was treated for the first 7 days at the Center for Clinical Veterinary Medicine at the LMU in Munich. Treatment groups were blinded until day 7 of the study. The cats remained in their owners' homes for the remaining days of the study and returned every 2 weeks for follow-up examinations and diagnostic tests at the clinic. Tests included abdominal and thoracic ultrasonography, blood chemistry, hematology, urinalysis, measurement of viral RNA in effusion, blood, and feces, and anti-FCoV antibodies. Detailed cardiological and neurological examinations were performed at study entry. The final re-examination was performed 168 days after the start of treatment.
GS-441524 was supplied as 50mg tablets and was legally imported from the UK. Owners kept diaries documenting items such as activity, stool consistency, food intake and body weight. 19 cats (of 20) in each treatment group completed treatment. Two cats were euthanized during treatment (days 3 and 31) due to secondary complications.
Clinical remission was observed between days 14 and 84 with a median of 28 days, and within the first 42 days 37/40 cats went into complete clinical remission. Every cat that completed treatment showed significant improvement in hematological and clinical chemistry parameters. At the beginning of the study, viral RNA was detected in the blood of 35/40 cats, and by day 28 no more viremia was noted in any cat. During the second phase (days 42 to 84) of the study, in which only the long-term treatment group received the drugs, no significant differences were found in viral load in blood, effusion and feces or anti-FCOV antibodies. By 168 days, all 38 cats remaining in the study were in complete remission. Two cats with neurological or ocular signs also fully recovered during treatment.
The most frequently observed adverse events were diarrhea in 25/40 cats (20 % of which were diagnosed as severe based on stool evaluation), elevation of liver enzymes (mild to moderate) in 24/40 cats between days 1 and 84, lymphocytosis in 27/40 cats and a slight increase in SDMA above the reference interval in 25/40 cats. None of the patients experienced adverse effects related to the administration of GS-441524.
This study demonstrated that a shorter treatment of 42 days with oral GS-441524 was as effective as the currently recommended 84-day treatment. GS-441524 was generally well tolerated, with no significant adverse reactions noted. Limitations include that all patients received continuous professional veterinary care during the first 7 days of treatment, which is not common in most clinical practice. In addition, only patients with wet FIP were included and only the oral form of GS-441524 was used. The preparation used in the study was legally manufactured in a strictly controlled manner by BOVA Specials in London, UK. Many cat owners around the world still purchase oral and/or injectable GS-441524 from "black market" sources, so it is unknown whether the 42-day treatment is equally effective in these patients. -BJP
More details:
Pedersen NC, Perron M, Bannasch M, Montgomery E, Murakami E, Liepnieks M, Liu H. Efficacy and safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis.J Feline Med Surg. 2019 Apr;21(4):271-281. doi: 10.1177/1098612X19825701. Epub 2019 Feb 13. PMID: 30755068; PMCID: PMC6435921.
Murphy BG, Perron M, Murakami E, Bauer K, Park Y, Eckstrand C, Liepnieks M, Pedersen NC. The nucleoside analog GS-441524 strongly inhibits feline infectious peritonitis (FIP) virus in tissue culture and experimental cat infection studies.Vet Microbiol. 2018 Jun; 219:226-233.doi: 10.1016/j.vetmic.2018.04.026. Epub 2018 Apr 22. PMID: 29778200; PMCID: PMC7117434.
When discussing feline coronavirus (FCoV) infection in a multi-cat environment, it is important to understand the correct nomenclature. The term FCoV is a collective term for two historically named viruses. The coronavirus was eventually identified as the causative agent of feline infectious peritonitis (FIP) in cats and named FIP virus or FIPV (Ward, 1970; Zook et al., 1968). FIPV was subsequently found to be a mutant form of FCoV that was present in cats infected with the widespread and minimally pathogenic enteric coronavirus and was named feline enteric coronavirus (FECV) (Pedersen et al., 1981). To avoid misunderstanding, this author prefers to refer to the form of FCoV that is relevant to the discussion. Therefore, it is appropriate to use the term FIPV when discussing the form of FCoV that is found in a specific type of white blood cell (monocyte/macrophage) in affected tissues and body fluids of cats with FIP. The term FECV is used when referring to the form of FCoV that causes chronic and intermittent infections of the epithelium in the lower intestine of healthy cats and is shed in large quantities in the feces. Enzootic is the correct term for infections that are maintained at a low and variable level in an animal population, while endemic is the corresponding term used for humans. Epizootic refers to a sudden and significant outbreak of a new infection, usually with rapid direct spread to animals of all ages. The human equivalent of epizootic is epidemic. Clinical “signs” are what veterinarians and doctors observe during a physical examination or what owners/parents convey to them, while symptoms are what people recognize in themselves and tell their doctors about.
FECV, like other feline mucosal pathogens, is maintained in the population as a persistent or recurrent latent infection (ie, enzootic). FECV is first shed in faeces from around 9–10 weeks of life, coinciding with the loss of maternal immunity (Pedersen et al., 2008). Infection occurs via the faecal-oral route and targets the intestinal epithelium, and the primary symptoms of enteritis are mild or mild, transient and rarely chronic or severe (Pedersen et al., 2008; Vogel et al., 2010). Subsequent excretion of feces is from the large intestine and usually stops after several weeks or months (Herrewegh et al., 1997; Pedersen et al., 2008; Vogel et al., 2010) with the development of immunity. The resulting immunity is unfortunately short-lived and repeated infections are common (Pearson et al., 2016; Pedersen et al., 2008. A stronger immunity appears to develop over time and cats older than 3 years appear to be less prone to re-infection and become with faecal excreters (Addie et al., 2003).
FIP is caused by specific FECV mutants that develop during infection (Poland et al., 1996; Vennema et al., 1995).1 A final risk factor for FIP in multifeline settings is the proportion of cats with high titers of antibodies to feline coronavirus and shedding virus in feces (Foley et al., 1997). FIP-causing mutants develop in 10 % or more cases of FECV infection, but only a fraction of these eventually cause disease (Poland et al., 1996). The actual incidence of FIP in a population with enzootic FECV infection appears to range from about 1% to 10% cats, with cases occurring at unpredictable intervals and varying from individual cases to small groups (Addie et al., 1995b; Foley et al. , 1997). The actual incidence appears to be driven by multiple host and environmental factors that somehow compromise the immune system and increase the risk of FIP.1
Given the direct relationship between the presence of FECV and FIP, a logical way to prevent FIP would be to minimize exposure to FECV. A vaccine would be the simplest approach to control FECV infection, but no vaccine can produce better immunity than recovery from natural infection, as demonstrated by the SARS-CoV-II vaccine (Li et al., 2019). Based on what is known about the weakness and short-lived nature of natural immunity to FECV (Pearson et al., 2016; Pedersen et al., 2008), together with the considerable variation in serotypes and strains between different populations and regions (Addie et al., 1995b; Liu et al., 2019), it is unlikely that effective vaccines against FECV will be developed.
Although enzootic FECV infection cannot be easily prevented by vaccination, it is possible to eliminate FECV from a closed group of cats through thorough carrier testing and strict quarantine (Hickman et al., 1995). However, FECV is so ubiquitous in nature and easily spread through direct and indirect cat-to-cat contact and on human-borne fomites that the strictest quarantine facilities and procedures are required to stop it. How strict must the quarantine be? Experience with testing and removal in conjunction with quarantine to eliminate and prevent FECV infection is limited to one report (Hickman et al., 1995). FECV was eliminated from a specific pathogen-free breed of cats at UC Davis by removing the virus shedders and significantly tightening quarantine procedures for the remaining colony (Hickman et al., 1995). Nevertheless, FECV re-entered this colony after several years, despite all attempts to prevent its spread (Pedersen NC, UC Davis, unpublished, 2022). The only example of effective quarantine for FECV was described for cats in the Falkland Islands (Addie et al., 2012). These islands in the remote South Atlantic have fortunately remained free of FECV, probably due to their extreme isolation. Measures have been taken to prevent future inadvertent introduction of FECV to the islands (Addie et al., 2012). Based on this experience, it is unlikely that FECV could be kept out of any group of domesticated cats unless the strictest isolation and infection prevention practices are followed.
An interesting approach to preventing or delaying FECV infection in kittens in catteries has been termed “early weaning and isolation” (Addie et al., 1995a). This approach was based on the observation that kittens born to mothers exposed to or infected with FECV acquired maternal immunity to infection by 9 weeks of age (Pedersen et al., 2008). Therefore, kittens weaned several weeks before the loss of this immunity (4–6 weeks of age) are usually free of infection and, if removed from their mothers and isolated from other cats, could theoretically be maintained free of the virus. This approach was initially popular, but the facilities and quarantine procedures required to prevent the introduction of the virus are difficult to maintain in catteries with larger numbers of breeding cats (≥ 5 dams, Hartmann et al., 2005). Therefore, elimination of FECV in kittens by early weaning and isolation has been doomed to failure in most conventional homes/catteries due to the continued direct and indirect exposure of infected cats to FECV. Another problem with early weaning and isolation is the need to separate virus-free kittens from other cats in a large group. This problem could be circumvented if all cats were cleared of infection at the same time. This can be achieved by serially testing feces for FECV excretion over a period of time and culling all excreting cats, along with strict quarantine. However, since a significant proportion of cats in households involved in enzootic FECV disease excrete feces (Foley et al., 1997; Herrewegh et al., 1997), elimination of cats can have a serious impact on the gene pool (Hickman et al., 1995). This begs the question – is there a way to eliminate FECV in all cats in a group at the same time?
Interestingly, the relatively recent discovery of effective antivirals against FIP (Pedersen et al., 2018, 2019) has also provided a theoretical method to eliminate all shedding viruses at once. Initial studies on this use of antivirals, although relatively preliminary, have suggested that FECV can be eliminated from a closed group of cats with a relatively short course of treatment (Addie et al., 2023). Assuming that FECV can be eliminated as an enzootic virus from a group of cats using specific antivirals, what are the pitfalls of such an approach? The first pitfall concerns the duration of immunity to reinfection that a short course of antivirals might induce. A follow-up study of cats successfully treated for FIP with GS441524 showed a return of low FECV shedding in 5/18 individuals within 3 to 12 months (Zwicklbauer et al., 2023), suggesting that treatment, like recovery from natural infection, does not confer long-term immunity. The second challenge is the cost of antivirals to treat primary and secondary infections, frequent stool testing to monitor shedding, and the establishment and maintenance of adequate quarantine facilities and practices. Therefore, home facilities with weak barrier isolation procedures are doomed to failure to keep this group of cats free of FECV for extended periods. The third challenge relates to the routine activities associated with breeding and exhibiting breeding cats. Breeding breeding cats involves frequent interaction between kittens and older cats, as well as between humans in contact with the cats and each other. It is also difficult to imagine that a breeder of purebred cats and an avid cat showgoer would forgo all the joys of breeding and showing their cats by avoiding all such contact. The ultimate question is – “now that the cats are free of FECV, what should be done with them?” What are the chances that they will remain free of FECV for any length of time after leaving the controlled environment? They will have no immunity to FECV and will be very sensitive to even the slightest exposure. The same will be true for the group of cats from which they come. Finally, and this is the biggest concern, the constant antiviral treatment required to keep a group of cats free of FECV infection will cause drug resistance to develop. We now know that resistance to GS-441524 can occur in cats treated for FIP, and researchers at UC Davis1 and Cornell University3 agree that acquisition of drug resistance in enzootic FECV infections would outweigh any potential benefit of such treatment on FIP incidence. FIP is currently curable in more than 90 % cases3, and even if resistance to antivirals does develop, it is largely confined to the affected cat. Arguably, HIV-1 infection in humans is currently prevented by antivirals, with no reported concerns about drug resistance. However, HIV-1 prevention treatment is not a monotherapy, but includes several drugs of different classes. Treatment with multiple drugs is not carried out with the aim of increasing the effectiveness of the treatment, but rather with the aim of preventing the development of drug resistance. If the virus develops resistance to one drug in the drug cocktail, the other drugs will prevent it from replicating.
In conclusion, I would like to paraphrase: “Just because something can be done, should it be done?” The author believes that much larger and better designed studies, conducted over a long period of time, are needed before treating asymptomatic FECV infection with antivirals is seriously considered as a means of preventing FIP. The overall incidence of FIP in smaller, well-maintained kennels, shelters and research breeding colonies with enzootic FECV infection is often low, and currently more than 90% of the FIP cases that could arise can be cured (Pedersen et al, 2019).3 A practical way to reduce the incidence of FIP is to keep the number of breeding cats and kittens low, to keep a larger number of older cats, not to breed individuals and bloodlines that have given rise to cases of FIP, and to minimize the stress of frequent introductions of new cats and placement/relocation .1 Isolation and early weaning can also be useful in smaller farms (Addie et al., 1995a). The problem of FIP in foster/rescue facilities is a bigger problem because most cats come from the feral population and are often very young when they arrive. They often suffer from malnutrition, a number of other diseases and are exposed to a high degree of stress associated with capture, routine treatment, change of diet, adaptation to a new environment and finally rehoming.1,3
References
Addie DD, Bellini F, Covell-Ritchie J, Crowe B, Curran S, Fosbery M, Hills S, Johnson E, Johnson C, Lloyd S, Jarrett O. 2023. Stopping Feline Coronavirus Shedding Prevents Feline Infectious Peritonitis. Viruses. 15(4), 818.
Addie DD, Schaap IA, Nicolson L, Jarrett O, 2003. Persistence and transmission of natural type I feline coronavirus infection. J Gen Virol. 84, 2735–2744.
Addie, D.; Jarrett, O. Control of feline coronavirus infections in breeding catteries by serotesting, isolation, and early weaning. 1995a. Feline Pract. 23, 92–95.
Addie DD, Toth S, Murray GD, Jarrett O. 1995b. Risk of feline infectious peritonitis in cats naturally infected with feline coronavirus. Am J Vet Res. 56, 429-34.
Foley JE, Poland A, Carlson J, Pedersen NC, 1997. Risk factors for feline infectious peritonitis among cats in multiple-cat environments with endemic feline enteric coronavirus. J Amer Vet Med Assoc. 210, 13131318.
Hartmann K, 2005. Feline infectious peritonitis Vet Clin North Am Small Anim Pract. 35(1), 39– 79.
Herrewegh AAPM, Mähler M, Hedrich HJ, Haagmans BL, Egberink HF, Horzinek MC, Rottier PJM, de Groot RJ, 1997. Persistence and evolution of feline coronavirus in a closed cat-breeding colony. Virology 234, 349–363.
Hickman MA, Morris JG, Rogers QR, Pedersen NC, 1995. Elimination of feline coronavirus infection from a large experimental specific pathogen-free cat breeding colony by serologic testing and isolation, Feline Practice 23, 96–102.
Li C, Liu Q, Kong F, Guo D, Zhai J, Su M, Sun D. 2019. Circulation and genetic diversity of Feline coronavirus type I and II from clinically healthy and FIP-suspected cats in China. Transbound Emerg Dis. 66, 763-775.
Pearson M, LaVoy A, Evans S, Vilander A, Webb C, Graham B, Musselman E, LeCureux J, VandeWoude S, Dean GA, 2019. Mucosal Immune Response to Feline Enteric Coronavirus Infection. Viruses 11, 906.
Pedersen NC, Theilen G, Keane MA, Fairbanks L, Mason T, Orser B, Che CH, Allison C, 1977. Studies of naturally transmitted feline leukemia virus infection. Am J Vet Res. 38, 1523–1531.
Pedersen NC, Boyle JF, Floyd K, Fudge A, Barker J, 1981. An enteric coronavirus infection of cats and its relationship to feline infectious peritonitis. Am J Vet Res. 42, 368-377. 5
Pedersen NC, Allen CE, Lyons LA, 2008. Pathogenesis of feline enteric coronavirus infection. J Feline Med Surg. 10, 529–541.
Pedersen NC, Liu H, Dodd KA, Pesavento PA, 2009. Significance of coronavirus mutants in feces and diseased tissues of cats suffering from feline infectious peritonitis. Viruses 1, 166-184.
Pedersen NC, Kim Y, Liu H, Galasiti Kankanamalage AC, Eckstrand C, Groutas WC, Bannasch M, Meadows JM, Chang KO, 2018. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. J Feline Med Surg. 20, 378-392.
Pedersen NC, Perron M, Bannasch M, Montgomery E, Murakami E, Liepnieks M, Liu H, 2019. Efficacy and
safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis. J Feline Med Surg. 21, 271-281.
Poland AM, Vennema H, Foley JE, Pedersen NC, 1996. Two related strains of feline infectious peritonitis virus isolated from immunocompromised cats infected with the feline enteric coronavirus. J Clin Microbiol. 34, 3180-3184.
Uusküla A, Pisarev H, Tisler A., et al., 2023. Risk of SARS-CoV-2 infection and hospitalization in individuals with natural, vaccine-induced and hybrid immunity: a retrospective population-based cohort study from Estonia. Sci Rep 13, 20347.
Vennema H, Poland A, Foley J, Pedersen NC, 1995. Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology 243, 150–157.
Vogel L, Van der Lubben M, , Te Lintelo EG, Bekker CPJ, Geerts T, Schuif LS, Grinwis GCM, Egberink HF, Rottier PJM, 2010. Pathogenic characteristics of persistent feline enteric coronavirus infection in cats. Vet Res. 41, 71.
Ward JM, 1970. Morphogenesis of a virus in cats with experimental feline infectious peritonitis. Virology 41, 191–194.
Zook BC, King NW, Robinson RL, McCombs HL, 1968. Ultrastructural evidence for the viral etiology of feline infectious peritonitis. Vet Path. 5, 91–95.
Zwicklbauer K, Krentz D, Hartmann K, et al., 2023. Long-term follow-up of cats in complete remission after treatment of feline infectious peritonitis with oral GS-441524. J Feline Med Surg. 25(8)
Footnotes
Pedersen NC. History of Feline infectious Peritonitis 1963-2022 – First description to Successful Treatment. https://sockfip.org/wp-content/uploads/2022/04/Review-FIP-1963-2022-final-version.pdf4.29.22.pdf.
Cornell University blog. Fight FIP. Unraveling feline infectious peritonitis from the ground up. https://blogs.cornell.edu/fightfip/fip-antivirals/.
Between January and April 2023, the Urolite Center in Minnesota received three shipments of atypical stones (Figure 1). All three samples were obtained from cats. All three cats were under 1 year old. Cats came from North and South America. In each case, the infrared spectrographic pattern of the stones was identical. Urinary stones usually contain large amounts of phosphorus, calcium and magnesium. In these cases, electron dispersion spectroscopy revealed a high proportion of nitrogen, carbon and oxygen.
Mystery solved. When asked about their medical history, all three cats were diagnosed with feline infectious peritonitis. All three were treated with either Remdesivir or its metabolite GS-441524. We requested samples of their antiviral drugs for analysis. The antiviral drugs were spectrographically identical (Figure 2). The stones were composed of GS-441524.
Figure 2: FT-IR spectroscopy of urinary stone of patient and reference sample GS-441524
After administration, GS-441524 is excreted primarily in the urine. Although GS-441524 is very soluble in organic solvents such as DMSO (10-59 mg/ml), it is poorly soluble in aqueous solutions such as water (0.0004 to 0.1 mg/ml). Its limited solubility makes GS-441524 a prime candidate for stone formation. Observation of urinary symptoms in cats receiving Remdesivir or GS-441524 is an indication to look for stones. Observation of atypical crystalluria or uroliths may be an indication to limit the dose of the drug (if possible) and increase water consumption to minimize stone formation.
What is FIP? – FIP is caused by a common and mostly harmless enteric coronavirus, similar to those that cause the common cold in humans and diarrhea in foals, calves and poultry. Most cats are infected with feline enteric coronavirus (FECV) at around 9 weeks of age and may be reinfected before 3 years of age, when cycles of infection become less frequent. Specific mutations that allow FECV to escape from the cells lining the lower intestine and infect the most basic cell of the immune system, the macrophage, occur in about 10 % infections. However, this macrophage infection is eliminated in all but 0.3–1.4 % cats. Predisposing conditions that lead to disease in this small proportion of cats include young age, genetic susceptibility, sex, overcrowding, poor nutrition, and a number of stressful events in the environment. The site of initial onset of the disease is in the lymphoid tissue in the lower small intestine, cecum, and proximal colon. Infected macrophages leave these initial sites of disease and migrate locally and in the bloodstream to small veins in the lining of the peritoneal cavity, the uveal tract of the eye, the ependyma, and the meninges and spine. Symptoms of the disease appear within days, weeks, sometimes months, and rarely a year or longer. The form of the disease that manifests itself is simply referred to as wet (effusive) or dry (non-effusive). The two forms are easily distinguishable, although there may be intermediate forms between them. Some cats may have symptoms of dry FIP but later develop wet FIP, or vice versa. Overall, about two-thirds of cats have wet FIP and one-third have dry FIP. The duration of illness until death, usually by euthanasia, used to be only a matter of days or weeks. Fewer than 5 % diseased cats, especially those with milder forms of dry FIP, survive longer than one year with the best symptomatic care.
Manifestations and forms of FIP
Clinical manifestations of FIP – The clinical manifestations of wet (Table 1) and dry (Table 2) FIP differ depending on the site(s) in the body where the infected macrophages end up causing inflammation. The intensity and nature of the inflammation are responsible for the form of the disease. Wet FIP is a more acute and severe form of FIP and is characterized by the accumulation of inflammatory fluid in either the abdominal cavity and/or the chest cavity. Involvement of the central nervous system (CNS) and eyes is relatively rare in the wet form of FIP (Table 1). The dry form of FIP is not characterized by diffuse inflammation and fluid discharge, but rather by fewer and more tumor-like lesions (ie, granulomas) in organs (e.g., kidneys, cecum, colon, liver, lungs, lymph nodes) in the abdomen or chest cavity or in the eyes and brain (Table 2). While the brain and/or eyes are involved in only 9 % cases of the wet form, neurological and/or ocular disease is the main clinical sign in 70 % cats with the dry form of FIP.
TABLE 1. VARIABILITY OF CLINICAL SYMPTOMS OF THE EFFECTIVE (WET) FIP IN CATS AVOIDED AT UC DAVIS
Symptoms associated with:
occurrence (%)
Peritoneal cavity
58%
Peritoneal and pleural cavities
22%
Pleural cavity
11%
Peritoneal cavity, eyes
2,8%
Peritoneal cavity, CNS *
1,9%
Peritoneal and pleural cavity, CNS
0,9%
Peritoneal and pleural cavity, eyes
0,9%
Pleural cavity, CNS, eyes
0,9%
Peritoneal cavity, CNS, eyes
0,9%
* CNS - Central nervous system (brain, spine)
TABLE 2. VARIABILITY OF CLINICAL SYMPTOMS OF NON-FUSION (DRY) FIP IN CATS AVOIDED AT UC DAVIS
Symptoms associated with:
occurrence (%)
Peritoneal cavity
30%
CNS
22%
Eyes
14%
CNS and eyes
8%
Peritoneal cavity, eyes
7%
Peritoneal and pleural cavities
4%
Peritoneal and pleural cavity, CNS
3%
Peritoneal and pleural cavity, eyes
2%
Peritoneal cavity, CNS, eyes
2%
Pleural cavity
1%
Blood-brain and blood-eye barrier
Basic facts - The eye and central nervous system (CNS) are protected from harmful substances by blood-eye barriers (blood-eye barrier) and blood-brain (blood-brain barrier). These barriers are of great evolutionary importance because they protect brain and eye functions from the effects of systemic toxins and infectious agents. Such barriers have been developed over millions of years by positive selection of the most capable individuals. The blood-brain barrier in cats does not pass about 80% most drugs, while the blood-eye barrier about 70%. Therefore, if a given dose of a drug such as GS-441524 reaches an effective blood level (plasma) of 10 μM, the levels in the brain (cerebrospinal fluid) will be only 2 μM and the level in the eye (ventricular water) will only be 3 μM. However, higher levels are likely to be reached in inflamed tissues and will decrease as inflammation subsides. This may be one of the explanations for the rapid improvement that is often observed in the first days of treatment.
Several other aspects of these two blood barriers need to be considered. First, their impermeability of undesirable substances varies from individual to individual. Second, the effectiveness of this barrier decreases in inflamed tissues and increases as inflammation subsides. This is good for treatment in the early stages of the disease, but bad for treatment in the final stages when the inflammation disappears and only the virus remains. Third, there are no simple, safe or effective means of weakening these barriers, and the only way to increase the level of the drug in the brain or eyes is to increase their level in the blood plasma by administering a higher dose, either orally or parenterally.
How these barriers affect forms of FIP - Paradoxically, ocular and neurological forms of FIP are also a consequence of the same barriers, but in this case in neurological and / or ocular FIP, the main problem is the entry of antibodies and immune lymphocytes. The phenomenon of neurological disease after a common systemic viral infection is well known in humans and animals. A typical example is polio-encephalomyelitis in humans and canine distemper in dogs. Poliomyelitis virus (polio) is a common intestinal pathogen and usually causes a mild or mild intestinal infection. However, in some people, the virus also penetrates the brain and spinal cord. Humans develop a strong systemic immune response to the polio virus, which is highly effective in eliminating the virus in all parts of the body, except the nervous system, where the limits of the blood-brain barrier are an obstacle to immunity. These unfortunates develop a classic neurological form of infection. A similar phenomenon occurs in canine distemper. Canine distemper virus, which is closely related to the human measles virus, causes an acute respiratory infection in young dogs, which manifests 7-14 days after exposure and lasts one to two weeks. Most of these dogs recover completely, but some develop neurological disease in three or more weeks. This highly lethal secondary form of canine distemper is caused by a virus that has escaped from the body to the brain and spinal cord during the respiratory phase of the infection and is protected from the host's immune system by the blood-brain barrier.
The distribution of the disease between the CNS and other parts of the body may also explain why blood tests are rarely abnormal in cats with primary neurological disease or in those who have relapsed to these forms during or after treatment with non-neurological forms of FIP. It appears that inflammation at privileged sites such as the CNS may not elicit a systemic inflammatory response and may not cause significant changes in hematology, nor an increase in total protein and globulin, and a decrease in albumin to globulin A: G ratio.
Preliminary diagnosis of ocular and neurological FIP
Preliminary diagnosis – Eye and neurological diseases are much less common in cats with wet than with dry FIP (Tables 1, 2). They also occur in primary and secondary forms. Primary disease accounts for approximately one-third of cases of dry FIP (Table 2), and lesions outside the eyes and central nervous system (CNS) are either absent or not readily discernible. Secondary neurological and ocular forms of FIP become much more common as a result of antiviral therapy and occur either during the initial treatment of the common extra-ocular/CNS forms or as a relapse during the 12-week post-treatment observation period.
The initial suspicion of neurologic and/or ocular FIP is based on age, origin, and presenting clinical signs. FIP occurs mainly in cats under 7 years of age, three-quarters of them under 3 years of age and with the highest incidence between 16 weeks and 1.5 years. Common symptoms in both ocular and neurological FIP were stunted growth in kittens and adolescent cats, weight loss in adults, and vague signs of ill health often associated with fever.
It is believed that the diagnosis of FIP, especially the dry form, is difficult. However, a preliminary diagnosis is relatively easy to establish due to stereotypic signaling, clinical history and physical findings, and the rarity of disease confusion in the group with the highest risk of FIP. Neurological and/or ocular forms of FIP can be confused with systemic feline toxoplasmosis, so many cats with these forms of FIP are tested for toxoplasmosis and treated with clindamycin. However, systemic toxoplasmosis is an extremely rare disease in cats, especially compared to FIP. FIP is easily distinguished by the cat's origin (breeding station, foster/rescue station, shelter), signaling (age, sex, breed) and basic blood test results. Deep fungal infections (coccidioidomycosis, blastomycosis, histoplasmosis) can cause ocular and sometimes neurological symptoms similar to FIP, but are still rare even in their endemic areas. Lymphoma can also be a differential diagnosis of dry FIP, but this disease is usually sporadic and occurs in older cats. A number of congenital disorders can also present with progressive neurological signs, but these occur mainly in younger cats and are not associated with the inflammatory manifestations of infectious diseases such as FIP, toxoplasmosis or deep mycoses.
Symptoms of ocular FIP - Ocular disease occurs as the sole or primary symptom in about one-third of cats with dry FIP and in two-thirds of cases associated with extra ocular lesions (Table 2). Eye disease is an unusual manifestation in cats that initially had wet FIP (Table 1). The initial clinical manifestation is unilateral or bilateral anterior uveitis, manifested by a change in iris color, turbidity and remnants of flocculant in the anterior chamber, keratic clots on the back of the cornea, and anisocoria (unequal pupil size). In some cats, retinitis (inflammation of the retina) is an accompanying feature, and is manifested by focal wallpaper hyporeflectivity associated with local inflammation and microhemorrhage (minor bleeding) of the retinal vessels. Less than one-third of cats with ocular FIP also show indeterminate or overt neurological symptoms (Table 2). In some cases, glaucoma, usually unilateral, and panopthalmlitis (inflammation of all layers of the eye) occur, which can lead to enucleation (removal of the eye).
Symptoms of neurological FIP - the same prodromal signs have often been observed in cats with neurological signs, but include vague signs of dementia, aggressive behavior, compulsive licking of inanimate objects and other cats, reluctance to jump to high places, spontaneous muscle twitching, abnormal swallowing movements and occasional seizures. Later symptoms include posterior ataxia, inability to jump to high places, physical and auditory hyperesthesia, hyperreflexia, and cerebellar-vestibular signs (cruciate extensor reflex, loss of conscious proprioception), seizures, and increasing incoordination and dementia. Symptoms of spinal involvement often include fecal and/or urinary incontinence, paralysis of the tail and hind limbs, pain in the lower back. Catastrophic decerebral symptoms are also associated with sudden and severe herniation of the brain into the spinal cord.
Confirmatory tests of ocular and neurological FIP
Basic facts - The definitive diagnosis of FIP is based on the identification of the presence of viral antigen or RNA in macrophages in typical effusions or lesions by PCR or immunohistochemistry (IHC). Definitive diagnosis can be a difficult and expensive process in many cats, and PCR / IHC can be false negative in up to 30% samples. In most cases, however, it is not necessary to go that far because of the diagnosis. A comprehensive set of historical, physical, and less direct laboratory abnormalities may be sufficient to make a diagnosis.
Laboratory symptoms - The diagnosis of ocular and neurological FIP can usually be made by combining characteristic changes in cerebrospinal fluid (CSF) and aqueous humor (high protein, high cell counts, neutrophils, lymphocytes, macrophages) with significant abnormalities in history and history, physical examination, CBC ), biochemistry, or MRI. Total protein concentration is often increased (mean, 9.4 g / l; median 3.6 g / l; range 0.85-28.8 g / l) as well as increased erythroblast (NRBC) count (mean 196 / μL median 171 / μL; range 15–479 / μL). Neutrophils are the dominant inflammatory cell in most cats, while lymphocytes and a mixture of neutrophils and lymphocytes are observed in a smaller proportion.
MRI is a useful tool for diagnosing neurological FIP, especially in combination with routine signaling / history, typical clinical signs, and CSF analysis. MRI identified three different clinical syndromes in 24 cats with an autopsy confirmed by neurological FIP (Rissi DR, JVDI, 2018.30: 392–399): 1) T3-L3 myelopathy, 2) central vestibular syndrome, and 3) multifocal CNS disease. In all cases, MRI abnormalities were found, including increased meningeal contrast, increased ependymal contrast, ventriculomegaly, syringomyelia, and foramen magnum herniation. 15 cases showed hydrocephalus (10 cases), cerebellar herniation through the foramen magnum (6 cases), swelling of the brain with flattened gyri (2 cases) and fibrin accumulation in the ventricles (2 cases) or leptomening (1 case). Histologically, 3 main different distributions of neuropathological changes were observed, namely periventricular encephalitis (12 cases), rombencephalitis (8 cases) and diffuse leptomeningitis with superficial encephalitis (6 cases).
In one study, the most useful anti-mortem indicator of neurological FIP was the positive titer of IgG anti-coronavirus antibodies in the CSF. Antibody titers in CSF 1: 640 or higher were found only in cats with FIP and RT-PCR was always positive. Initial studies indicated that the antibody present in the CSF was produced, at least in part, in the CNS. However, in another study, the antibody was detected only in cats with serum titers of 1: 4096 to 1: 16384, and the researchers concluded that the antibodies in the CSF were obtained passively. In another attempt to measure local CNS antibody production in cats with FIP, the albumin quotient and IgG index were measured to determine if the proteins in the CSF were of blood origin or of local origin. Neither the albumin quotient nor the IgG index identified a pattern consistent with intrathecal IgG synthesis in cats with the CNS form of FIP. In conclusion, anti-coronavirus antibodies appear to enter the CSF at high levels, when they are also at high serum levels. Indeed, IFA serum coronavirus antibody titers in cats with ocular and neurological FIP are among the highest in any form of FIP.
PCR test performed from a sample of CSF and aqueous humor with a higher number of proteins and cells is highly sensitive and specific for ocular and neurological FIP. However, it is recommended that only a PCR test targeting the FCoV 7b gene be used, and no less sensitive PCR to FIPV specific mutations in the S gene. This FCoV gene is often used for PCR because it is the most abundant viral transcript and is therefore likely to that it will be detected. In some PCR assays, the FCoV M gene was targeted because it is highly conserved in all isolates, but transcripts are less numerous than in the 7b gene.
Immunohistochemistry on cells collected from spinal fluid is as sensitive and specific as PCR on samples with higher protein and cell counts. The antigen is localized specifically to macrophage-like cells.
The rapid FIP response to GS-441524 is being used as a confirmatory test increasingly. However, it should only be used in cases where there is other supporting evidence for a diagnosis of FIP. However, the truth is that there are probably no other simpler or cheaper means available at the moment to facilitate the diagnosis.
Treatment of neurological and ocular FIP
Difficulties in obtaining authorization for veterinary use of medicinal products for human use – Pharmaceutical companies such as Gilead Sciences and Merck have refused to compromise the development and approval processes of their promising anti-coronavirus drugs such as GS-5734 (Veklury®/Remdesivir) and EIDD-2801 (Molnupiravir®) or their respective biologically active forms GS-441524 and EIDD -1931. Out of desperation, cat owners around the world have turned to the Chinese black market for drugs like GS-441524. This black market was not entirely motivated by profit – China's FIP problem also grew at the same time as the domestic cat population. Moreover, even if Gilead Sciences had approved the use of GS-441524 in animals, the immediate need for an effective treatment for FIP has overtaken the official approval and commercialization process, which takes many years. Chemical companies and a dozen or more vendors of injectable and oral products have been able to satisfy the demand for GS from tens of thousands of desperate cat owners around the world. Veterinarians have been reluctant to pressure human pharmaceutical companies like Gilead to license their promising antiviral drugs for use in animals, but they are increasingly involved in helping owners with treatments. It therefore appears that the unapproved use of human drugs such as GS-441524, which are also desperately needed in veterinary species, will be the norm for many years to come.
(This paragraph comes from the original article from 1/4/2021.)
Virus-specific inhibitors – Inhibition of viral genes regulating specific stages of infection and replication has become the mainstay of treatment for chronic RNA virus infections in humans, such as HIV and hepatitis C virus. Currently, two classes of antiviral drugs have been shown to be effective against FIP. The first class consists of RNA synthesis inhibitors and includes the nucleoside analogs GS-441524 (the active ingredient in Remdesvir) and EIDD-2801 (molnupiravir). The second class of drugs consists of viral protease inhibitors, such as GC376 (prodrug of GC373) and Nirmatrelvir (prodrug of nitrile modification of GC373). Protease inhibitors are much less effective at crossing the blood-brain and blood-ocular barriers than nucleoside analogues and are not recommended for the treatment of neurological or ocular FIP.
Treatment with GS-441524 – GS-441524 has become the drug of first choice for the treatment of cats with all forms of FIP, and both injectable (SC) and oral forms are available in the off-label Chinese market. However, oral absorption is less than 50 % effective compared to injection, thus requiring twice the dosage of oral GS-441524. Suppliers of oral GS-441524 almost never disclose the actual concentration of GS-441524 in tablets or capsules, but rather label them as an equivalent injection dose. There is also an upper limit to the absorption efficiency of oral GS, making it difficult to achieve the higher blood levels needed to reach sufficient amounts of the drug in the brain and eyes. Therefore, if cats with ocular and neurological disease fail despite high equivalent doses of oral GS-441524, a switch to injectable GS-441524 should be considered before switching to a drug such as molnupiravir is considered.
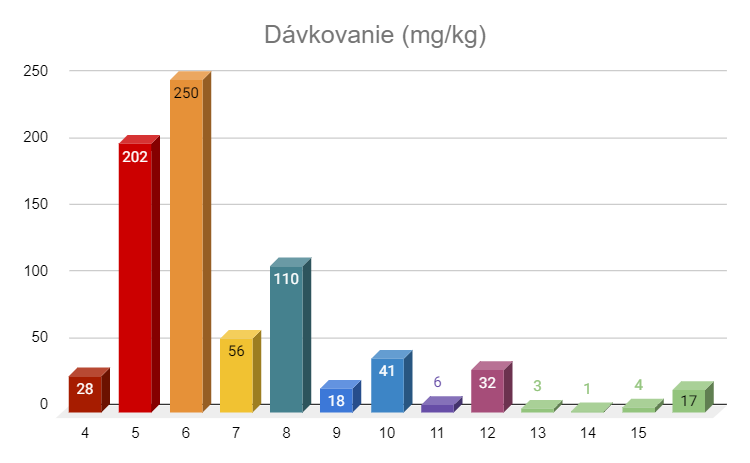
The starting dose for cats with wet or dry FIP and no signs of ocular or neurological disease is 4-6 mg/kg daily for 12 weeks, with younger and wet cases tending towards the lower end and dry cases towards the higher end. Cats with eye lesions and no neurological signs are started at 8 mg/kg daily for 12 weeks. Cats with neurological signs are started at 10 mg/kg daily for 12 weeks. If cats with wet or dry FIP initially develop ocular or neurological signs, they are switched to the appropriate ocular or neurological doses. The dose of GS is adjusted weekly to account for weight gain. Weight gain can be huge in many of these cats, either because they are in poor condition to begin with or because their growth has been stunted. If the cat does not gain weight during treatment, this is considered a bad sign. The initial dosage is not changed unless there are serious reasons for this, such as ineffectiveness of treatment or improvement in blood test values, improvement is very slow, low activity level, initial clinical symptoms have not resolved, or the disease form has changed with the appearance of ocular or neurological symptoms. If there are good reasons to increase the dosage, it should always be from +2 to +5 mg/kg per day and for at least 4 weeks. If these 4 weeks exceed the original 12-week treatment time, the treatment time is extended. A positive response to any increase in dosage can be expected, and if you don't see an improvement, it means that the dosage is still not high enough, drug resistance is emerging, the GS mark is not what it should be, the cat does not have FIP, or there are other diseases that confuse the treatment.
One of the most difficult decisions is determining when to stop treatment. Although some cats, often younger with wet FIP, can be cured as early as 8 weeks and possibly earlier, the usual treatment period is 12 weeks. Some cats may even require dose adjustments and even longer treatment periods. Critical blood levels such as hematocrit, total protein, albumin and globulin levels and absolute lymphocyte counts usually return to normal in curing cats after 8 to 10 weeks, when there is often an unexpected increase in activity levels. We believe, but there is no evidence yet, that after 8-10 weeks, the cat will have its own immune response against the infection. This is a situation that occurs in the treatment of people with hepatitis C, which is also a chronic RNA virus infection that often requires antiviral treatment for up to 12 weeks or more.
Cats with ocular disease and no neurological impairment show a rapid response to GS, and complete recovery of vision with minimal or no residual damage is expected in as little as two weeks. Cats that develop neurological abnormalities, develop neurological disease during the treatment of other forms of FIP, or develop neurological symptoms during the 12-week post-treatment observation period also improve rapidly, but the dose is much higher, the duration of treatment often longer and the cure rate slightly lower. Treatment failures in cats with neurological FIP are due to either insufficient dose or the development of drug resistance.
Unfortunately, there is no simple blood test that can determine when a cat with neurological impairment has fully recovered. Many cats with neurologic FIP show minimal blood abnormalities, especially those with primary neurologic FIP, and the abnormalities often disappear by the end of treatment, even though residual sites of inflammation remain in the brain or spinal cord. In addition, some cats that recover from the infection will have mild to moderate neurological deficits that are residual effects of the previous illness. These facts make it difficult to use blood test results or residual neurological deficits as indicators of cure or undertreatment. Although a thorough eye examination can clearly rule out active signs of disease, the true state of the disease in the brain and spinal cord can only be determined by an MRI, ideally together with an analysis of the cerebrospinal fluid. These procedures are expensive, not available to everyone, and may not provide definitive proof that the infection in the CNS has been cleared.
Fear of relapses means that many people involved in GS treatment are too cautious about a single blood parameter that is slightly abnormal (eg, slightly high globulin or slightly low A: G ratio), or final ultrasound results suggesting suspiciously enlarged abdominals. lymph nodes, small amounts of abdominal fluid or blurred irregularities in organs such as the kidneys, spleen, pancreas or intestines. It should be borne in mind that the normal range of blood values applies to most animals, but it is a bell-shaped curve, and that there are a few non-standard patients who will have values at the edge of these curves. Ultrasonographers must consider the degree of pathology that can occur in the FIP of the affected abdomen and how scars and other permanent consequences can change the normal appearance of successfully treated cats. In situations where such questions arise, it is better to focus in more detail on the overall picture and not just on one small part. The most important outcome of treatment is a return to normal health, which has two components - external signs of health and internal signs of health. External signs of health include a return to normal activity levels, an appetite, adequate weight gain or growth, and coat quality. The latter is one of the best criteria for cat health. Internal health symptoms include the return of certain critical values to normal based on periodic complete blood count (CBC) monitoring and serum chemical profiles. The most important values in CBC are hematocrit and relative and absolute total white blood cell, neutrophil and lymphocyte counts. The most important serum values for chemical analysis (or serum electrophoresis) are total protein, globulin, albumin and A: G ratio levels. Bilirubin is often elevated in cats with effusive FIP and may be useful in monitoring the severity and duration of inflammation. There are many other values in the CBC panels and serum, and it is not uncommon for some of them to be slightly higher or lower than normal, and it is better to ignore these values unless they are significantly elevated and associated with clinical signs. For example, high BUN and creatinine, which is also associated with increased water consumption, excessive urination, and urinary abnormalities. The number of machine-counted platelets in cats is notoriously low due to the trauma of blood collection and platelet aggregation and should always be verified by manual examination of blood smears. The final decision to discontinue or extend treatment when faced with unclear doubts about different testing procedures should always be based on external manifestations of health than on any single test result.
(This paragraph comes from the original article from 1/4/2021.)
Relapses usually refer to infections that have escaped into the central nervous system (brain, spine, eyes) during treatment for wet or dry FIP that are not accompanied by neurological or ocular symptoms. Doses of GS-441524 used to treat these forms of FIP are often insufficient to effectively cross the blood-brain or blood-ocular barrier. The blood-brain barrier is even more efficient than the blood-ocular barrier, which explains why eye lesions are easier to heal than brain and/or spinal cord infections. Post-treatment relapses involving the eyes, brain, or spine are usually treated for at least 8 weeks at an initial daily dose at least 5 mg/kg higher than the dose used during primary treatment (eg, 10, 12, 15 mg/kg per day). Cats that fail to clear the infection at doses up to 15 mg/kg per day are likely to have developed varying degrees of resistance to GS-441524. Partial resistance may allow suppression of disease symptoms but not cure, while complete resistance is manifested by varying severity of clinical symptoms during treatment.
Different groups focused on the treatment of FIP have made various modifications in the treatment protocols. Some groups will treat with an extremely high dose of GS from the beginning and not increase the dose when indicated, or will recommend discontinuing or extending the high dose for the last two weeks in the hope that this will reduce the risk of relapse. In addition to GS, systemic prednisolone is often prescribed, but should only be used temporarily to stabilize serious illness. Systemic steroids reduce inflammation but tend to mask the beneficial effects of GS, and if used for an unreasonably long time and in high doses, can interfere with the development of immunity to FIP. Restoration of immunity to FIP is thought to be an important part of successful GS treatment. Therefore, some people advocate the use of interferon omega or non-specific immunostimulants to further stimulate the immune system, and some come up with other modifications. There is no evidence that using an extremely high dose will improve the cure rate. Also, interferon omega and non-specific immunostimulants have not been shown to have beneficial effects on FIP, whether given as a single treatment or as an adjunct to GS. The practice of adding another antiviral drug, the viral protease inhibitor GC376, to the treatment of GS in cats that develop resistance to GS is also emerging, but this still requires further research. Finally, it is common for owners, treatment groups and veterinarians to add many supplements, tonics or injections (eg B12) to increase hematopoiesis or to prevent liver or kidney disease. However, such supplements are rarely necessary in cats with pure FIP.
Molnupiravir (EIDD-2801) – Molnupiravir is very similar to GS-441524, but is a cytidine rather than an adenine nucleoside analog. It is widely used as an oral treatment for early cases of COVID-19 in humans, but in the last 1-2 years it has been increasingly used to treat cats with FIP. Due to the toxicity observed in cats at higher doses and as yet unknown chronic side effects, it is most often recommended for cats that developed resistance to GS-441524 during primary treatment or relapsed with neurological/ocular signs after treatment with high doses of GS- 441524. Fortunately, molnupiravir has a different resistance profile than GS-441524.
The safe and effective dosing of molnupiravir in cats with FIP has not been established in properly controlled and monitored field studies such as those performed for GC376 and GS-441524. However, the estimated starting dose of molnupiravir in cats with FIP was derived from published EIDD-1931 and EIDD-2801 in vitro cell culture studies and other laboratory and experimental animal studies. Molnupiravir (EIDD-2801) has an EC50 of 0.4 µM/µL against FIPV in cell culture, while the EC50 of GS-441524 is approximately 1.0 µM/µL. Molnupiravir begins to show cellular cytotoxicity at concentrations of 400 µM or higher, while GS-441524 is non-toxic at 400 µM. Both have similar oral absorption of around 40-50 %. The current recommended starting dose of molnupiravir for neurologic and ocular FIP is 8–10 mg/kg orally every 12 hours for 84 days. Depending on the response to treatment, it may be necessary to increase it to a maximum of 15 mg/kg orally every 12 hours. At higher doses, molnupiravir toxicity is likely to occur as indicated by changes in the complete blood count.
Causes of treatment failure
Incorrect dosage adjustments - It is important to start treatment with the appropriate dosage and to monitor it closely with regular checks on temperature, weight and external signs of improvement. The CBC and serum chemical analysis panel, which contains baseline protein values (total protein, albumin, globulin (TP - albumin = globulin) and A: G), should be performed at least once a month. with GS-441524 Expensive serum protein electrophoresis does not provide much more valuable information.
Low quality GS-441524 - GS-441524 is not approved for marketing in any country and is sourced from a small number of Chinese chemical companies which sell it to distributors as pure powder. Vendors dilute it into injectable solutions or prepare oral forms for sale under their trade names. There is no independent mechanism to ensure the quality of the final product sold to cat owners. Nevertheless, the main providers of dilute forms for injectable solutions and / or oral preparations are surprisingly honest, and some even offer limited guarantees if treatment with some of their products does not cure the disease. However, the batches sold by some providers appear to be counterfeit and some are not in the specified concentration. There may also be differences between batches, probably due to occasional problems with the supply of raw GS by retailers or problems with meeting the needs and expectations of the cat owner. Various groups of FIP Warriors have good information about the most reliable brands.
Drug resistance - resistance to GS-441524 may already exist at the time of diagnosis, but this is unusual. It occurs more frequently during treatment and is initially only partial and requires only higher doses. In some cats, it may become complete. Resistance is the biggest problem in cats with neurological disease, or they develop brain infections during treatment or within a few days or weeks after stopping treatment. Many cats with partial drug resistance may be "treated" for their symptoms, but they relapse as soon as treatment is stopped, as is the case with HIV treatment, for example. There are cats that have been able to partially or completely treat the symptoms of FIP for more than a year, but without a cure. Resistance eventually worsens and the symptoms of the disease worsen, treatment difficulties become unbearable for the owner or the owner's financial resources run out.
GS side effects
GS-441524 treatment is incredibly free of systemic side effects. It can cause mild kidney damage in cats without significant kidney damage, but does not lead to latent disease or kidney failure. Systemic drug reactions such as vasculitis have been observed in several cats and can be confused with injection site reactions. However, these drug reactions are in places where injections are not given, and often stop on their own or respond well to short-term low-dose steroids. The main side effect of GS treatment is pain at the injection sites, which varies from cat to cat and according to the abilities of the person giving the injections (usually the owner). Swelling or ulcers at the injection site sometimes occur in owners who do not change the application site often enough (do not stay between the shoulder blades) and do not inject into the muscular and nervous layers under the skin. I recommend choosing places starting one inch behind the shoulder blades, down from the back to 1 to 2 inches in front of the root of the tail and one third to half way down to the chest and abdomen. Many people use gabapentin to relieve pain before injections. Swollen spots and ulcers at the injection site should be stripped of surrounding hair and gently cleaned 4 or more times a day with sterile cotton swabs soaked in homemade hydrogen peroxide diluted 1: 5. They usually do not require any more complicated treatment and will heal in about 2 weeks.
Prognosis of treatment with GS441524
Exact cure rate data with GS-441524 are not yet available, but it seems possible to cure more than 80% cats with confirmed FIP. Treatment failure is due to misdiagnosis of FIP, inadequate treatment monitoring and dose adjustment, complicating diseases, poor GS, resistance to GS, or economic difficulties. The cure rate is slightly lower in cats with neurological forms of FIP and in older cats. Older cats are more susceptible to other chronic diseases, which either predispose cats to FIP or complicate overall health.
Cats with neurological FIP may suffer permanent residual symptoms of the disease. This is especially true for cats with spinal involvement and urinary and/or fecal incontinence or hind paralysis. Hydrocephalus and syringomyelia are common complications of neurological FIP and often persist to some extent after the infection has cleared. Fortunately, most cats with neurologic FIP recover normal or near-normal function despite persistent traces of hydrocephalus and syringomyelia.
Legal treatment for FIP?
We hope that the legal form GS-441524 will be available soon. The drug, called Remdesivir, is the greatest hope of the present because Remdsivir breaks down into GS when given intravenously to humans, mice, primates and cats. Remdesivir (Veklury®) has been fully approved by the US FDA and similar approval is likely to follow in other countries. If so, it can be prescribed by any licensed human physician as well as veterinarians. However, the use of Remdesivir in the United States was initially limited to a specific subset of patients with Covid-19 and only under controlled conditions and with ongoing data collection. Until all restrictions are lifted, it will not be easily accessible for human use. We have no experience in treating cats with Remdesivir instead of GS-441524. The molar basis of Remdesivir is theoretically the same as GS-441524. GS-441524 has a molecular weight of 291.3 g / M, while Remdesivir is 442.3 g / M. Therefore, 442.3 / 291.3 = 1.5 mg of Remdesivir would be required to obtain 1 mg of GS-441524. The diluent for Remdesivir is significantly different from the diluent used for GS-441524 and intended for intravenous use in humans. How diluted Remdesivir will behave when given by subcutaneous injection over 12 weeks or more is not known. Mild signs of hepatic and renal toxicity were observed in humans. GS-441524 causes mild and progressive renal toxicity in cats, but without apparent hepatic toxicity. It is uncertain whether renal toxicity observed in humans receiving Remdesivir is due to its active substance (ie GS-441524) or to chemical agents designed to increase antiviral activity.
The GC376 approval process for cats (and humans) is ongoing at Anivive, but will take two or more years. GC376 is a viral protease inhibitor and, unlike GS-441524, which inhibits the initial stage of viral RNA replication, GC376 prevents viral replication in the final stage of its replication process. Therefore, it is unlikely to have a significant synergistic viral inhibitory effect and its use in combination will be much more important in inhibiting drug resistance (e.g. in combination antiviral therapy for HIV / AIDS).
Improper use of GS-441524
Some veterinarians, in collaboration with major Chinese supplier GS-441524, have advocated its use to eliminate feline enteric coronavirus (FECV) infection. The reason is to prevent the occurrence of a mutant virus causing FIP (FIPV) and thus prevent FIP. This approach was supported by limited and highly controversial studies with shelter cats, which were naturally exposed to the FECV. Although this approach is attractive at first glance, it is a very incorrect use of GS-441524 in cats. FECV infection originally occurs in kittens and is not associated with any significant symptoms of the disease. Elimination lasts for weeks, months, and in some cases indefinitely, but in most cats, it eventually stops when immunity develops. Most cats over the age of three will no longer shed the virus. GS-441524 treatment is highly unlikely to result in more permanent immunity than is observed in nature and to eliminate cycles of infection and reinfection in younger cats.
Although our current knowledge of FECV infection seriously challenges this approach, there are even more compelling reasons why we will not treat healthy cats GS-441525 or other antiviral agents in the future. We already know from published studies that some primary strains of FIPV are resistant to GS-441524 (and GC376). We also know that drug resistance has become a long-term problem in cats with long-term treatment for GS-441524, especially in neurological forms of FIP. Therefore, the use of drugs such as GS-441524 in a large population of healthy cats will undoubtedly lead to widespread resistance to enzootic FECV. This resistance will also manifest itself in FIP-causing FECV (FIPV) mutations from these populations, making it impossible to use GS-441524 in more and more FIP cats. Unfortunately, veterinary medicine does not have the means of human medicine, it is not stimulated by potential benefits, which would lead to the discovery, testing and approval of more and more antiviral drugs to circumvent either natural or acquired drug resistance, which is already the case in HIV / AIDS treatment. achieved (at least on time).
(This part comes from the original article from 1/4/2021.)
Niels C. Pedersen, DVM PhD Distinguished Professor Emeritus UC Davis, Center for Animal Health Companion January 4, 2021, updated February 10, 2023
dr. Sam Taylor BVetMed(Hons) CertSAM DipECVIM-CA MANZCVS FRCVS Prof. Séverine Tasker BVSc BSc DSAM PhD DipECVIM-CA FHEA FRCVS, Prof. Danielle Gunn-Moore BSc(Hon), BVM&S, PhD, MANZCVS, FHEA, FRSB, FRCVS Dr. Emi Barker BSc BVSc PhD PGCertTLHE DipECVIM-CA MRCVS, Dr. Stephanie Sorrell BVetMed(Hons) MANZCVS DipECVIM-CA MRCVS
Given the current situation, Sam Taylor, Séverine Tasker, Danièlle Gunn-Moore, Emi Barker and Stephanie Sorrell discuss treatment protocols to help doctors manage this viral disease.
Introduction
Figure 1: Remdesivir intended for intravenous or subcutaneous administration
In August 2021, remdesivir (Figure 1) became legally available to UK vets to treat FIP in cats. Since then, many cats and kittens have been and are still being successfully treated. As with any new product, protocol modifications are adopted with experience, and in light of the recent release (November 2021) of oral GS-441524 (50 mg tablets) from a specialist UK manufacturer (Figure 2), this article has been drafted to support general practitioners in the use of remdesivir and GS-441524 in the treatment of FIP. It should be kept in mind that treatment may need to be tailored to the individual cat based on the client's response, compatibility and financial capabilities. The specific protocols below may help veterinarians and their clients, but will not be appropriate for all cases.
Treatment protocols (updated November 2021)
The dosage of the drugs has been increased compared to previous recommendations based on the experience of our Australian colleagues, who have treated more than 600 cats so far. Although some cats responded to previously recommended lower doses, they found that relapse was possible at or near the end of the 84-day (12-week) treatment period, leading to the need to extend treatment with a higher daily dosage. This was ultimately more expensive than starting treatment at a higher dosage.
Figure 2: GS-441524 oral tablets
With the use of remdesivir and/or GS-441524, treatment options are now available including a 12-week course of injectable remdesivir, switching from injectable remdesivir to oral GS-441524, or an exclusively oral GS-441524 protocol.
Suggested dosing, benefits, and limitations of each protocol are listed below. Remdesivir cannot be taken orally. The recommended dosage of drugs (Table 1) depends on the clinical picture - ie whether there is an effusion or not and whether there is eye and/or neurological involvement - this is due to differences in drug penetration into tissues. In case of doubt, it is more appropriate to use a higher dosage.
Please note that these dosages of oral GS-441524 are higher than reported in some publications - this is because these publications used so-called black market preparations of GS-441524 in which the amount of active ingredient administered to cats was not confirmed. The dosages given in this article are based on experience using an oral formulation of known GS-441524 that is legally available in the UK and Australia. Therefore, extrapolation cannot be used for other oral preparations for which the active ingredient and/or its concentration is not known or is not indicated by the manufacturer.
Combined injection and oral treatment protocols
The decision when to switch from injectable remdesivir to oral GS-441524 may depend on tolerability of injections (or oral tablets), differences in product cost (including cost of needles, syringes, sharps disposal, losses), owner preference, and finances.
Experience suggests that this transition may occur between days 7 and 14 after initiation of intravenous or subcutaneous remdesivir therapy. The change can be made directly; remdesivir is given for one day and GS tablets are started the next day.
The protocol chosen depends on the severity of the FIP disease in the cat. Dosage is shown in Table 1.
Serious condition
If the condition is severe (anorexia, dehydration, the cat is usually hospitalized):
Initial treatment with remdesivir given once daily intravenously (Table 1) for three to four days – ie days 1, 2, 3 and/or 4. This will achieve the loading dose of the drug. Each day, dilute the required dose of remdesivir to a total volume of 10 mL with saline and administer slowly over 20 to 30 minutes by hand or pump.
Subsequently, administer SC remdesivir once daily at the same dose (Table 1) until days 7 to 14.
On days 8-15, switch to oral GS-441524 once (or twice) daily (Table 1) and continue until at least day 84.
Table 1: Overview of dosing recommendations for remdesivir and GS-441524
Clinical presentation
Remdesivir - by injection
GS-441524 – oral
Cats with effusion and no ocular or neurological signs
10 mg/kg once a day
10-12 mg/kg once a day
No effusion and no ocular or neurological symptoms
12 mg/kg once a day
10-12 mg/kg once a day
Ocular symptoms present (effusive and non-effusive FIP)
15 mg/kg once a day
15 mg/kg once a day
Neurological symptoms (effusive and non-effusive FIP)
20 mg/kg once a day
10 mg / kg twice daily (ie 20 mg/kg in divided doses)
Translator's Note: the injectable form of GS-441524 is not used for legal treatment in the UK. Given that the molecular weight of remdesivir is approximately 2x higher than the molecular weight of GS-441524, the recommended dosage of remdesivir is approximately 2x higher than that of GS-441524. Coincidentally, the bioavailability of GS-441524 when administered orally is about 50%, so the dosage for tablets with the stated real GS content from the manufacturer BOVA is practically identical to the dosage for remdesivir injection.
Less serious condition
Regarding a less severe condition (normal hydration, food intake):
Initial treatment with remdesivir SC 1x a day (Table 1) until the 7th to the 14th day.
Change to 1x (or 2x if a very high neurological dose is required) daily oral administration of GS-441524 (Table 1) on days 8-15 and continue until at least day 84.
An exclusively oral protocol
In the event that injectable treatment is not tolerated/financially feasible, only the oral GS-441524 treatment protocol is recommended:
1x (or 2x if a very high neurological dose is required) daily oral GS-441524 (Table 1) for at least 84 days.
Possible side effects of remdesivir:
Remdesivir appears to be well tolerated. However, the following side effects have been reported:
Transient local discomfort/stinging on injection (see prevention later).
Development/worsening of a pleural effusion (not always proteinaceous) during the first 48 hours of treatment, sometimes requiring drainage.
Cats may be depressed or nauseous for several hours after intravenous administration.
An increase in the activity of the enzyme alanine aminotransferase has been reported (whether due to the underlying disease of FIP or an adverse effect of the drug is unclear).
Mild peripheral eosinophilia has been reported.
A note on weighing cats
During treatment, it is very important to weigh cats weekly using an accurate scale - with successful treatment, kittens will gain weight and/or grow, which will require a dose increase to ensure that the dose of antiviral given is still appropriate for the type of FIP being treated.
Options for clients with a limited budget
Please note that ideally, treatment should be administered using the recommended preparations and dosage for as long as possible (up to 84 days) to increase the likelihood of a cure.
Use the options below only when absolutely necessary, as a relapse may occur, which then requires longer treatment, leading to increased costs:
Administer oral treatment with GS-441524 only for 84 days as above.
Administer injectable remdesivir or oral GS-441524 for as many days as the owner can tolerate, then switch to oral mefloquine 62.5 mg two to three times weekly (in large cats three times weekly) or 20 mg to 25 mg orally once daily (if possible to change the composition of the tablets - for example, Novalabs) to complete the 84-day treatment protocol; mefloquine is less expensive than remdesivir and GS-441524, but further research is needed to assess its effectiveness in this setting.
If it is necessary to increase the dose of remdesivir (for example, due to a neurological disease that appears during treatment), but it is not possible to afford it, mefloquine treatment can be added as an adjunctive treatment, because it is cheaper than remdesivir, although it is necessary to assess the effect of this combination further research.
Feline interferon omega has also been used in the post-remdesivir/GS-441524 treatment period, but further research is needed to assess whether this combination is necessary.
Is the oral treatment given with or without food?
GS-441524 is administered on an empty stomach (with some water) - food may be administered 30 minutes after administration of the drug.
Mefloquine is given with food, otherwise vomiting often occurs.
Remember to support clients when giving oral medications, as this can also be challenging for them. Direct clients to the website iCatCare, where you can find information and videos.
How can I help owners with remdesivir SC application?
Remdesivir injection may cause temporary local discomfort. The following measures can help reduce discomfort and improve cooperation:
Make sure owners use a new needle each time to withdraw medication from the vial (this will reduce the risk of bacterial contamination of the vial, as well as rubbing the top of the reusable seal vial with alcohol before inserting the needle).
Make sure owners change the needle after removing the medicine from the bottle and before giving the injection (puncturing the reusable seal will blunt the needle).
Needle size preferences vary; some prefer a 21G needle to make the injection faster; others find the finer 23G needle better tolerated, so it may be worth trying both if you have problems.
Alternate injection sites.
Allow remdesivir to warm to room temperature before administration.
Oral gabapentin (50 mg to 100 mg per cat) and/or intramuscular or SC buprenorphine given at least 30 to 60 minutes before remdesivir injection may be useful to induce mild sedation/analgesia.
The area to be injected may also be shaved to help owners locate a suitable injection site and to allow topical EMLA cream to be applied 40 minutes prior to injection, although superficial desensitization may not help as discomfort is usually caused by remdesivir under the skin.
Ensure that the full injection dose is always administered and encourage owners to report any failures as this may influence decisions in case of relapse.
Cats will need several weeks of treatment. Encourage owners to make the injection more enjoyable by using treats at the time of the injection or by petting, combing or playing with the cat if it is less motivated to eat. Suggest that owners spend time with their cat in a positive way each day to avoid any damage to the cat-owner relationship that may reduce cooperation.
What can I expect during treatment?
During the first two to five days, you should see an improvement in behavior, appetite, resolution of pyrexia, and a decrease in abdominal (Figure 3) or pleural fluid if an effusion is present (please note that in some cases pleural fluid may be transient in the first few days worsen - if the cat is at home, advise the owner to measure the resting respiratory rate and respiratory effort) - the effusion usually subsides within two weeks.
If discharge is still present after two weeks, consider increasing the dosage.
Serum albumin increases and globulin decreases (that is, normalizes) within one to three weeks, but note that globulins may initially increase when a large volume of effusion is absorbed.
The resolution of lymphopenia and anemia may take longer, up to 10 weeks.
Mild peripheral eosinophilia is a common finding and may be a favorable marker for disease resolution, similar to that seen in patients with COVID-19.
The size of the lymph nodes will decrease within a few weeks.
If progress is not as expected, consider reassessing the diagnosis (see below) and/or increasing the dosage.
Figure 2. Cat with FIP and ascites. Effusions should begin to subside within three to five days of starting treatment.
What should be observed during treatment?
Ideally, serum biochemistry and hematology after two weeks and monthly thereafter.
For clients on a limited budget, monitor only weight/behavior/effusion/neurological signs/key biochemical abnormalities (for example, measuring only globulin and bilirubin).
Note that the activity of the enzyme alanine transaminase (ALT) may increase - it is not clear whether this is due to the pathology of FIP or a reaction to the drug, and it is not usually a reason to stop treatment. It is not known whether the addition of hepatoprotective treatment (eg S-adenosyl-L-methionine) helps in these cases.
Ultrasonography in the outpatient clinic to monitor the resolution of the effusion and/or the size of the lymph nodes.
If I observe a positive response to the treatment, when should I stop the treatment?
Not earlier than after 84 days (12 weeks).
Verify the disappearance of previous abnormalities (clinical, sono, biochemical and hematological examination).
Discontinue treatment only after the cat has been normal (clinically, biochemically and hematologically) for at least two weeks (ideally four weeks).
What should I do if I have no or only a partial response to treatment?
Make sure the cat actually has FIP - reevaluate the diagnosis, look for other pathologies, consider repeated sampling (eg, external laboratory analysis of any fluid; cytology or lymph node biopsy).
If biochemical abnormalities (especially hyperglobulinemia and albumin to globulin ratio) remain present after 6 to 8 weeks, increase the dosage as for relapse (see below) by 3 mg/kg to 5 mg/kg daily and continue treatment, not stopping until parameters have normalized for at least 2 weeks, as indicated above under “when to stop treatment?” – This may mean extending treatment beyond 12 weeks.
What should I monitor after treatment?
Advise the owner to monitor the cat closely for recurrence of the clinical condition - this monitoring should continue for 12 weeks after the end of treatment.
Ideally, repeat serum biochemistry and hematology two weeks and one month after stopping treatment (to detect any changes that might indicate an early relapse).
Note that relapse may occur with clinical symptoms but without any significant biochemical/hematological abnormalities.
Relapse
In case of relapse – e.g. recurrence of effusion, pyrexia, development of ocular or neurological symptoms, or return of hyperglobulinemia:
Make sure the cat has FIP - reassess the diagnosis, consider other pathologies, consider repeat sampling (for example, external laboratory analysis of any fluid, cytology or lymph node biopsy).
If relapse occurs during treatment, increase the dose of remdesivir or GS-441524 and monitor treatment as before, making sure that treatment is not stopped before the cat has been normal for at least two weeks. The increased dosage depends on the dosage the cat is receiving at the time of the relapse, the nature of the relapse and the financial possibilities, but can be up to the recommended dosage for neurological FIP (see above).
If relapse occurs after stopping treatment, restart remdesivir or GS-441524 at a higher dose (usually 3 mg/kg to 5 mg/kg daily higher than previously used doses) and continue treatment for an additional 12 weeks. The increased dosage used depends on the dosage the cat was receiving at the time of the relapse and the nature (eg severity and/or development of neurological signs) of the relapse, but may be up to the dosage recommended for neurological FIP (20 mg/kg - see Table 1). It is possible that some cats will respond to a shorter treatment, but ideally relapse treatment is continued for the full 12 weeks after treatment has been completed to prevent relapse.
If the dose of remdesivir or GS-441524 cannot be increased (for example, the highest neurologic dose of 20 mg/kg is already being used), consider mefloquine as adjunctive therapy (see above) while continuing remdesivir or GS-441524 at the same dose.
Castration and routine measures during the treatment of FIP
If the cat responds to treatment, neutering should ideally be done one month after the treatment has been completed. However, if leaving the cat unneutered causes significant stress – for example, escape attempts or stress when the mother is in heat – it is advisable to neuter during treatment. If the latter option is required, neutering should ideally be done when the cat is responding well to treatment and has at least two weeks of treatment remaining after neutering (so that the antiviral treatment is given during the potential “stress” of post-neutering).
There is no contraindication for routine deworming and flea treatment in cats treated with remdesivir or GS-441524.
No information is available on vaccination of cats treated for FIP. If the cat is well during treatment, it should be vaccinated as usual, as it is still likely that the vaccination will have a protective effect. For cats that have completed the initial round, consider giving a third dose of vaccine after completing FIP treatment (see WSAVA Vaccination Guidelines).
If veterinary procedures are required, the clinic stay should be minimized and protocols and handling should be implemented according to Cat Friendly Clinicto avoid stressing the cat.
Complementary treatment
If a cat is receiving prednisolone, it should be discontinued during administration of remdesivir or GS-441524, and then discontinued completely, unless needed for short-term treatment of a specific immune-mediated disease resulting from FIP—for example, hemolytic anemia.
Supportive therapy such as antiemetics, appetite stimulants, fluid therapy, and analgesics may be given along with remdesivir or GS-442415 as needed.
Possible future updates
We are constantly learning during treatment with these drugs, and recommendations may change over time. Other substances have been tested in cats, such as protease inhibitors (such as GC376) and other nucleoside analogues (such as molpurinavir), but these are not currently commercially available. How these agents and other immunomodulatory agents (such as polyprenyl immunostimulant) will fit into future protocols is currently unknown.
Translator's Note: The original article was published and updated in February 2022, since molnupiravir officially became available for the treatment of COVID-19 in humans, and there is also the possibility of its use in the treatment of FIP.
Acknowledgement
We thank Richard Malik and Sally Coggins for their advice in the preparation of this article.
dr. Richard Malik DVSc MVetClinStud PhD FASM graduated from the University of Sydney in 1981. He is a specialist in small animal internal medicine with a special interest in infectious diseases of dogs and cats. She works at the Center for Veterinary Education and helps organize CPD.
dr. Sally Coggins BVSc (hons I) MANZCVS (Feline Medicine) she graduated from the University of Sydney in 2007 with first class honours. Sally is currently investigating novel antiviral therapeutics for the treatment of feline infectious peritonitis and is conducting clinical trials open to national recruitment.
FIP advisory line
The above experts have come together to launch an email address ‘FIP advice’ (fipadvice@gmail.com) where they volunteer to answer questions about the new treatment and spread the word to vets and veterinary nurses in the UK. They have so far answered over 150 emails on the advice line.
Meagan Roy 1, Nicole Jacque 2, Wendy Novicoff 3, Emma Li 1,Rosa Negash 1 , Samantha JM Evans 1 *
Department of Veterinary Biosciences, College of Veterinary Medicine, The Ohio State University, Columbus, OH 43210, USA
Independent Researcher, San Jose, CA 95123, USA
Departments of Orthopedic Surgery and Public Health Sciences, School of Medicine, University of Virginia, Charlottesville, VA 22903, USA
* Author to whom correspondence should be addressed.
Academic editors: Alessia Giordano and Stefania Lauzi Pathogens 2022, 11(10), 1209; https://doi.org/10.3390/pathogens11101209 Received: 19/09/2022 / Revised: 9/10/2022 / Received: 19/10/2022 / Published: 20/10/2022 (This article is part of a special issue of Advances on Feline Coronavirus Infection)
Feline infectious peritonitis (FIP) is a complex and historically fatal disease, although recent advances in antiviral therapy have revealed treatment options. A newer therapeutic option, unlicensed molnupiravir, is used as first-line therapy for suspected FIP and as salvage therapy for cats that have persistent or recurrent clinical signs of FIP after treatment with GS-441524 and/or GC376. Treatment protocols for 30 cats were documented based on owner-reported data. 26 cats treated with unlicensed molnupiravir as rescue therapy were treated with a mean starting dose of 12.8 mg/kg and a mean final dose of 14.7 mg/kg twice daily for a median period of 12 weeks (IQR = 10-15). A total of 24 of the 26 cats were still living without signs of disease at the time of writing this report. One cat was euthanized after treatment due to persistent seizures and the other cat underwent retreatment due to relapse of clinical signs. Few adverse effects have been reported, with the most prominent - drooping ears (1), broken whiskers (1) and severe leukopenia (1) - occurring at doses above 23 mg/kg twice daily. This study provides proof of principle for the use of molnupiravir in cats and supports the need for future studies to further evaluate molnupiravir as a potentially safe and effective therapy for FIP.
Keywords: FIP; coronavirus; antiviral drug; EIDD-2801; black market
1. Introduction
Feline infectious peritonitis (FIP) is a complex and historically fatal disease caused by mutation of the ubiquitous feline enteric coronavirus (FECV) [1]. Recent advances in feline and antiviral medicine have revealed potential treatment options for FIP. The 3C-like protease inhibitor GC376 was the first targeted antiviral therapy used against this disease [2]. GC376 was highly effective in improving clinical signs of FIP in 19 of 20 naturally infected cats, but showed limited ability to manage long-term disease [2]. Pedersen et al. continued to investigate the antiviral compound GS-441524, a nucleoside analog and active metabolite of remdesivir (GS-5734). GS-441524 demonstrated superior ability to treat and control disease in naturally infected cats compared to GC-376, with 25 of 31 cats disease-free at the time of writing [3]. Since these discoveries, cat owners worldwide have obtained these mostly unlicensed drugs to treat their FIP cats with remarkably high success rates [4]. Legal FIP treatment is in high demand in the United States due to ethical and legal concerns regarding the unlicensed drugs GC376 and GS-441524. In addition, some cats with FIP have exhausted all current treatment options due to disease relapse and/or treatment failure after GS-441524, GC376 and/or combination therapy. Therefore, an effective and legal treatment option for FIP is urgently needed. In connection with the recent outbreak of SARS-CoV-2, a number of new antivirals have entered the market. Molnupiravir (EIDD-2801), manufactured by Merck, is currently available under an emergency use authorization (EUA) from the FDA for the treatment of COVID-19 in adults [5]. It is an oral prodrug of the nucleoside analog BD-N4-hydroxycytidine, which increases guanine to adenine and cytosine to uracil nucleotide transition mutations in coronaviruses [6]. This mechanism increases the rate of mutations above the accepted limit, which in turn inactivates the virus [7]. Molnupiravir has been found to be safe and well tolerated at doses up to 800 mg twice daily in patients with COVID-19 [8]. Some studies have reported significant reductions in hospitalizations and deaths in mild-to-moderate COVID-19 patients, although efficacy appears to be lacking in severe COVID-19 patients [7].
Because of molnupiravir's strong potential to treat other coronavirus infections, cat owners have begun using unlicensed molnupiravir (or its active metabolite EIDD-1931) purchased over the Internet to treat FIP. However, the use of molnupiravir for the treatment of FIP is currently not documented in any scientific literature. Unlicensed molnupiravir can be used as first-line therapy for suspected FIP, but also as rescue therapy to treat cats that have persistent or recurrent clinical signs of FIP after GS-441524 and/or GC376 therapy. The aim of this study is to document this use and provide proof of principle for molnupiravir as a potential treatment for FIP based on owner-reported data.
2. Materials and methods
The survey was conducted using the Qualtrics XM program (Qualtrics Version May-August 2022, Provo, UT, USA) under license from Ohio State University. The survey (Supplementary Data S1) was written in English and consisted of 94 multiple-choice and free-response questions asking about FIP diagnosis, clinical signs, initial therapy (used before molnupiravir), molnupiravir treatment, adverse events, duration of treatment, and remission time. The number of free-response questions was limited to limit recall bias. The survey also allowed owners to upload relevant documents (eg veterinary medical records and laboratory results). The survey was formatted using questions from previous studies to maintain consistency of language and style, as well as newly developed questions specific to the experience of molnupivir treatment. The logic of the survey dictated that some questions appeared only after a particular answer was selected, while others were skipped when a particular answer was selected. This conditional logic was used to reduce questionnaire completion bias and questionnaire fatigue. The survey took approximately 20-30 minutes to complete and could be saved and completed later if needed. This study was approved by the Ohio State University Institutional Review Board (Protocol No. 2021E0162).
The survey was distributed to participants individually by email and data were collected from June to August 2022. Participants were selected from a subset of owners seeking molnupiravir therapy for their cat with suspected FIP through popular FIP therapy and social media support groups. Inclusion criteria were surveys of cats suspected of having FIP based on veterinary diagnosis, failure to respond to initial therapy, or recurrence of clinical signs after completion of initial therapy other than molnupiravir (eg, GS-441524 or GC376) and completion of 8–10 weeks of oral molnupiravir therapy (or those who subsequently died or were euthanized during therapy). This study also included a small group of cats that received molnupiravir for 8-10 weeks as initial and sole therapy, which will be referred to as first-line therapy in the rest of this paper, when FIP is suspected. Exclusion criteria were surveys with incomplete data or cats not diagnosed with FIP by a veterinarian.
3. Results
3.1 Demographic data
A total of 80 potential participants were identified through a social media FIP support group and 37 invitations to complete the questionnaire were sent to those participants with available contact details. A total of 33 questionnaires were mailed and 21 participants were sent follow-up emails to obtain complete questionnaire data. Seventeen owners attached relevant documents to the submitted questionnaires and two additional owners sent relevant documents containing veterinary medical records, laboratory results and diagnostic images to the study email address. These submitted documents were used to document adverse reactions reported by one participant. One response was a refusal to participate. Two cases were excluded because the cats did not have a diagnosis of FIP from a veterinarian (one of them was reported to have been diagnosed based on the loss of a sibling due to FIP and the other was examined by a veterinarian who concluded that blood tests were not consistent with FIP). A total of 30 cats with suspected FIP were therefore included in this study, 4 of which had not received any treatment prior to molnupiravir administration. These four cats were included as a separate small cohort of first-line molnupiravir treatment. A flow chart of these cases is shown in Figure 1. The countries of origin represented were the United States (25), Germany (2), Poland (2), and Sweden (1). The sex/neuter status of the cats at the time of diagnosis was 40 % neutered males, 40 % spayed females, and 20 % intact males. The mean age at diagnosis was 9.7 months, with a range of 1 month to 6 years. The majority of cats were of mixed or unknown breed (70 %); These included seven purebred cats and two special crossbreeds (e.g., a Balinese-Ragdoll-Siamese cross). Responses that referred to the cat as "American Shorthair" or "American Longhair" were instead categorized as mixed breed, given the commonly reported confusion among American owners regarding the nomenclature of this breed.
Figure 1. This flowchart represents the number of cases in each treatment block.
Regarding comorbidities, feline leukemia virus was reported in only one cat and calicivirus was reported in one cat. Several cats also had a history of external and/or internal parasitic infections (3), conjunctivitis/ocular infections (2), and bacterial skin infections (pyoderma) (1). A total of 16 cats had neurological signs of FIP. Three cats had both neurological and ocular manifestations of FIP, and two cats had only ocular manifestations of FIP. Of the remaining cases, seven were effusive, while five cases were non-effusive. The full breakdown of FIP types is shown in Table 1.
Cat
Age at diagnosis (months)
Sex/castration status at diagnosis
Tribe
Previous medical conditions
Country of origin
FIP form
Duration of initial treatment (weeks)
Disease-free period
Second therapy
Duration of the second therapy (weeks)
Disease-free period
The third therapy
Duration of the third therapy (weeks)
Disease-free period
1
4
Male
European shorthair
parasitic infections, URI at an early age
Germany
neurological
injectable oral GS-441524
8
none
injectable and oral GS-441524
15
none
2
15
neutered cat
Burmese
none
Sweden
effusive, non-effusive, neurological
injectable GS-441524
12
less than 4 weeks
injectable GS-441524
14
17 days
oral GS-441524
5 weeks
none
3
9
neutered cat
British Shorthair
none
Poland
effusive, neurological, ocular
injectable GS-441524
13
less than 2 weeks
injectable GS-441524
12
more than 6 months, less than 1 year
4
5
neutered cat
Abyssinia
none
USA
effusive
injectable GS-441524
12
less than 2 weeks
injectable GS-441524
14
less than 4 weeks
5
4
neutered cat
Balinese/Ragdol mix
calicivirus, conjunctivitis, giardiasis, tapeworm, URI
USA
non-effusive
injectable GS-441524
13
less than 8 weeks
6
7
neutered cat
Siamese
none
USA
neurological
injectable and oral GS-441524, injectable GC, injectable and oral molnupiravir
12
none
7
7
neutered cat
American Shorthair
none
USA
non-effusive
injectable and oral GS-441524
5
none
8
6
neutered cat
American Shorthair/Siamese mix
tapeworm, FCoV
USA
effusive, neurological
injectable and oral GS-441524
5
none
9
4
neutered cat
Homemade mixed
broken pelvis
USA
effusive
injectable and oral GS-441524
14
less than 6 months
oral GS-441524
13
less than 4 weeks
oral GS-441524/injectable GC
6 weeks in combination then 6 weeks of oral GS
none
10
4
neutered cat
Homemade mixed
none
USA
effusive
injectable GS-441524
23
less than 4 weeks
11
72
neutered cat
Homemade mixed
FeLV
USA
non-effusive
oral GS-441524
12
less than 6 months
12
5
Male
Homemade mixed
none
USA
non-effusive, neurological, ocular
injectable and oral GS-441524
17
none
13
01.V
Male
Savannah
none
USA
effusive, neurological
injectable and oral GS-441524
24
less than 6 months
injectable and oral GS-441524
12
less than 4 weeks
14
4
neutered cat
Homemade mixed
Skin and eye infections, fleas
Poland
non-effusive, neurological
injectable GS-441524
12
less than 2 weeks
injectable GS-441524
17
less than 4 weeks
15
12
neutered cat
American Shorthair
none
USA
effusive
injectable GS-441524/GC
01.V
none
16
5
neutered cat
Homemade mixed
none
USA
effusive, neurological
injectable GS-441524
12
less than 4 weeks
17
4
Male
American longhair
none
USA
ocular
injectable and oral GS-441524, GC376
13
none
18
6
neutered cat
Homemade mixed
none
USA
effusive
injectable GS-441524
12
none
19
12
neutered cat
Homemade mixed
none
USA
non-effusive
injectable and oral GS-441524
12
less than 2 weeks
injectable GS-441524
12
none
20
6
neutered cat
Unknown
none
USA
non-effusive, neurological
injectable GS-441524
4
none
oral GS-441524
3
none
21
4
neutered cat
Norwegian forest
none
USA
neurological
injectable GS-441524
12
less than 6 months
injectable GS-441524
01.V
none
Molnupiravir, GS-441524, GC
12 weeks
none
22
6
neutered cat
Homemade mixed
none
USA
neurological, ocular
oral GS-441524
3
none
23
12
neutered cat
Unknown
none
Germany
neurological
injectable GS-441524
16
less than 6 months
24
3
Male
Homemade mixed
none
USA
neurological
injectable GS-441524
12
less than 6 months
25
6
neutered cat
American Shorthair
none
USA
effusive
oral GS-441524
13
less than 1 week
26
1
Male
Unknown
none
USA
non-effusive
injectable GS-441524
12
less than 1 week
27
7
neutered cat
Homemade mixed
none
USA
non-effusive, neurological
Molnupiravir
12
less than 1 week
*Molnupiravir
28
24
neutered cat
Homemade mixed
none
USA
effusive
Molnupiravir
29
12
neutered cat
Homemade mixed
none
USA
non-effusive, ocular
Molnupiravir
30
24
neutered cat
Homemade mixed
none
USA
neurological
Molnupiravir
Table 1. Signaling and initial therapy characteristics of all 30 cats treated with unlicensed molnupiravir for suspected FIP.
3.2. Initial treatment before initiation of molnupiravir
A total of 26 of 30 cats received initial treatment for suspected FIP with unlicensed GS-441524 or a drug combination containing unlicensed GS-441524 as the main base drug (GS-441524-based). Half (13) of the cats were treated with injectable GS-441524. Only three cats were treated with oral GS-441524, while the other seven cats were treated with a combination of injectable and oral GS-441524 throughout the treatment period. Two cats were treated with a combination of the unlicensed drug GS-441524 and the unlicensed drug GC376. Cube no. 6 was treated with all previously mentioned drugs along with molnupiravir for 12 weeks of a very complicated regimen (Supplementary Data S2). Dosing of combination drugs used as part of primary therapy (eg, GC376 and molnupiravir) was not determined. Reported initial doses of the unlicensed GS-441524 ranged from 2 mg/kg to 10 mg/kg; the most frequently reported dosages were 5-6 mg/kg (eight cats) and 10 mg/kg (seven cats). Most (21) cats received a dose once a day. Only four were dosed twice daily, and one cat was dosed twice daily for one week at first, then switched to once daily dosing. The median duration of treatment based on GS-441524 was 12 weeks (IQR = 12-13). In fifteen cats, a change in daily doses was reported during treatment. For several cats, the daily dose was increased by body weight to maintain the same dosage in mg/kg. Others increased the mg/kg dosage because of insufficient clinical response or a change in route of administration (eg, from injectable to oral GS-441524). No participant reported dose reduction during treatment.
A total of 6 of 26 cats completed a shorter than average 12-week treatment with GS-441524 due to insufficient clinical response and were immediately started on another treatment. Two of the six cats initiated a different route or dose of unlicensed GS-441524 treatment as shown in Table 1. One cat switched from injectable to oral GS-441524 treatment on the second treatment. In the second cat, the dose of GS-441524 was simply increased during the second treatment. The remaining four cats started treatment with unlicensed molnupiravir at this time, as shown in Table 2. Of the 20 cats that completed at least 12 weeks of treatment with GS-441524, 16 experienced clinical remission. All 16 were in remission for less than 6 months, with 2 cats in remission for less than a week before clinical signs returned. All 16 started a second round of treatment, with 10 receiving a second round of GS-441524-based treatment and 6 starting molnupiravir at this time. Four cats that completed treatment with GS-441524 but did not achieve clinical remission were immediately started on molnupivir. A total of 26 cats received primary treatment with GS-441524 and all 26 relapsed or did not respond adequately. A total of 10 of 26 completed a second round of GS-441524-based treatment and 16 started molnupivir treatment.
Cat
Clinical symptoms at the beginning of treatment
Brand name
Initial dosage and frequency
Final dosage and frequency
Duration of treatment (weeks)
Time to improve
Persistent clinical symptoms
The result
Adverse effects
1
diarrhea, vomiting
Aura Plus
11 mg/kg twice daily
11 mg/kg twice daily
12
less than 1 week
none
clinical remission
none
2
none reported
Aura
12 mg/kg twice a day
12 mg/kg twice a day
12
uncertain
none
clinical remission
none
3
anisocoria, colored spots in the eye, polydipsia, pica, weight loss
Aura 2801
28 mg/kg twice daily
14 mg/kg twice a day
12
within 2 weeks
none
clinical remission
none
4
anorexia, lethargy, weight loss
EIDD
7 mg/kg twice a day
7 mg/kg twice a day
12
less than 1 week
none
clinical remission
none
5
colored spots in the eye, diarrhea, hiding and lack of socialization
Aura 2801
6 mg/kg once daily
13 mg/kg once daily
10
within 2 weeks
none
clinical remission
none
6
anisocoria, constipation, anorexia, fecal and urinary incontinence, lethargy, paralysis, seizures, pale gums, weight loss
Aura 2801
20 mg/kg twice a day
20 mg/kg twice a day
11
less than 1 week
none
clinical remission
none
7
anorexia, difficulty walking, hiding, lack of socialization, jaundice, lethargy
anorexia, heavy walking, hiding, lack of socialization, lethargy, unusual timidity
Aura 2801
11 mg/kg twice daily
16 mg/kg twice a day
18
more than 4 weeks
nothing physical but the MRI is still not normal
clinical remission
none
15
blindness, constipation, anorexia, diarrhea, enlarged abdomen, hiding, lack of socialization, lethargy, pale gums, weight loss
Aura 2801
16 mg/kg twice a day
16 mg/kg twice a day
12
less than 1 week
none
clinical remission
none
16
anorexia, difficulty walking, lethargy, seizures, tremors, weight loss
Aura 2801
14 mg/kg twice a day
14 mg/kg twice a day
12
less than 1 week
none
clinical remission
none
17
cough, anorexia, difficulty breathing, hiding, lack of socialization, lethargy, vomiting, weight loss
Aura 2801 and Aura 1931
12 mg/kg twice a day
17 mg/kg twice a day
20
within 3 weeks
anorexia
clinical remission
nausea/vomiting, anorexia
18
constipation, anorexia, difficulty walking, hiding, lack of socialization, weight loss
Aura 2801
12 mg/kg twice a day
12 mg/kg twice a day
8
within 2 weeks
none
clinical remission
none
19
lethargy, anorexia
Aura 2801
12 mg/kg twice a day
12 mg/kg twice a day
7
within 2 weeks
none
clinical remission
none
20
trembling/shaking
Aura 2801
10 mg/kg twice a day
23 mg/kg two to three times a day
10
less than 1 week
in remission about 1 1 weeks before the onset of seizures
euthanasia
decreased appetite when dosed three times a day, severe leukopenia, loss of beard, scaly skin on ears
21
difficulty walking, fecal incontinence
Aura 2801 and Aura 1931
13 mg/kg twice a day
30 mg/kg twice a day
14
less than 1 week
difficult walking, difficult jumping, fecal incontinence persisted during the study (1 week post treatment)
relapse and euthanasia
drooping ear tips, muscle weakness
22
colored spots in the eye, anorexia, difficulty walking, hiding, lack of socialization, lethargy
Aura 2801
16 mg/kg twice a day
19 mg/kg twice a day
9
within 2 weeks
none
clinical remission
none
23
difficulty walking, anorexia, loss of balance
EIDD aura
12 mg/kg twice a day
15 mg/kg three times a day
10
within 2 weeks
heavy walking
clinical remission
none
24
blindness, colored spots in the mouth, anorexia, difficulty breathing, difficulty walking, enlarged abdomen, urinary incontinence, jaundice, lethargy, paralysis, tremors
Aura 2801
15 mg/kg twice a day
15 mg/kg twice a day
16
less than 1 week
none
clinical remission
none
25
difficulty breathing, difficulty walking, hiding, lack of socialization, lethargy, URI
Aura 2801
7 mg/kg twice a day
7 mg/kg twice a day
16
within 2 weeks
none
clinical remission
none
26
lethargy, anorexia
Aura 2801
14 mg/kg twice a day
14 mg/kg twice a day
15
less than 1 week
neurological twitches, elevated liver enzymes
clinical remission
none
Table 2. Treatment and outcome characteristics of 26 cats receiving unlicensed molnupiravir as rescue therapy.
3.3. The second round of treatment before the initiation of molnupiravir
Overall, 10 of 26 cats that received initial GS-441524 treatment and subsequently relapsed were reported to have received a second round of unlicensed GS-441524 treatment prior to initiation of molnupiravir. Again, most cats received injectable GS-441524 (6), with two receiving oral GS-441524 and two receiving both injectable and oral GS-441524. Reported dosages ranged from 4-5 mg/kg to 15 mg/kg; the most frequently used dosages were 7-8 mg/kg (two cats) and 15 mg/kg (two cats). Most cats were dosed once daily (seven cats), one cat was dosed twice daily and one cat was dosed three times daily. In most cats, the dose was varied during treatment. The two doses were weight-adjusted to maintain the same dosage in mg/kg. Dosing in mg/kg was increased in five cats that did not respond adequately or developed new clinical signs (eg, neurological signs). The median duration of treatment was 12.5 weeks (IQR 9.75–14.25). Only two cats did not undergo at least 12 weeks of therapy. One of the two added GC376 and molnupiravir to current GS-441524 therapy, and the other started molnupiravir as sole therapy. Of the eight cats that completed at least 12 weeks of GS-441524 therapy, two did not achieve clinical remission. Both cats started treatment with unlicensed molnupiravir at that time. The remaining six cats were reported to achieve clinical remission after a second round of treatment with GS-441524. Five of the six cats were in remission for less than 4 weeks, with the exception of one cat that was in remission for more than 6 months but less than a year. Seven out of ten cats started taking unlicensed molnupiravir at this time.
3.4. The third round of treatment before starting molnupiravir
The remaining three cats received a final round of GS-441524-based treatment before switching to molnupiravir. Cat no. 2 received oral GS-441524 for 5 weeks prior to initiation of molnupiravir. Cat no. 9 was treated for 6 weeks with oral and injectable GS-441524 and then continued for 6 weeks with oral GS-442524 alone. Dosing and frequency in both cats are unknown, as the survey collected data on only two therapies prior to molnupivir. Cat no. 21 received a combination of GS-441524, GC376 and molnupiravir for 12 weeks. The dosage, frequency and duration of each varied radically over the course of 12 weeks (Supplementary Data S3). All three cats started treatment with molnupiravir without clinical remission from this third round of treatment.
3.5. Molnupiravir as rescue therapy
Of the 26 cats receiving unlicensed molnupiravir as rescue therapy, most were using the Aura brand, with only 2 cats using a different brand of molnupiravir. More than 81 % cats (18) were treated with Aura 2801, 1 cat was treated with Aura 1931, and another 2 cats were treated with both Aura preparations. The mean initial dosage was 12.8 mg/kg twice daily. One cat was dosed only once a day and two cats were dosed 2 to 3 times a day. The most commonly used initial dosage was 12 mg/kg twice daily. Dosage ranged from 6 to 28 mg/kg twice daily. 11 dosage changes were reported, all but one being an increase in dosage. Reduction of dosage in cat no. 3 was not explained in any way. The mean final dosage was 14.7 mg/kg twice daily, with the same three cats differing in dosing frequency. The most common final dosage was also 12 mg/kg twice daily. The dosage range was 7 to 30 mg/kg twice daily.
Median duration of treatment was 12 weeks (IQR 10-15). Overall, a wide range of 7-20 weeks was reported. Only eight cats were treated for less than 12 weeks. A cat that completed only 7 weeks of treatment was reported to have discontinued treatment due to achieving clinical remission. All 26 cats completed treatment at 7 weeks or longer and all 26 cats survived. No cases of missed doses of molnupiravir have been reported.
Owners reported improvement in clinical signs in more than 92 % cats within three weeks of initiation of molnupiravir treatment, with 84.6 % cats showing improvement within two weeks and nearly half (46.2 %) within one week. Only two cases were reported differently, with one cat showing no signs of improvement for up to 1.5 months, and the owner of the other cat being unsure of the timescale and degree of improvement in clinical signs. A total of seven cats with persistent clinical signs of FIP were reported. In one of them, the disappearance of clinical symptoms was reported after one week of the observation period. Others are thought to have had residual symptoms such as difficulty walking or jumping, tremors, MRI changes and fecal incontinence. The full range of persistent clinical signs is shown in Table 2. Only three cats reported adverse reactions in response to molnupiravir, including nausea/vomiting, anorexia, drooping ear tips (Figure 2), brittle whiskers, leukopenia, scaly skin and muscle wasting. At the time of publication, 24 of 26 cats are living in clinical remission of FIP after oral molnupiravir treatment. One cat reportedly died 1 week after discontinuation of molnupiravir due to a prolonged seizure, and the other cat (No. 21) was disease-free 4 weeks before relapse. Cat no. 21 then started a second round of molnupiravir at the same dose, but was subsequently euthanized due to insufficient response to treatment.
Figure 2. Dropped ear tips were reported as an adverse effect of unlicensed molnupiravir treatment in cat no. 21.
In cat no. Severe leukopenia was reported in 22 cases. Through veterinary records, it was found that cat no. 22 has moderate panleukopenia with lymphopenia, neutropenia, and normal hem and thrombograms on 4 of 5 sequential complete blood counts, which were confirmed through veterinary records of sequential complete blood counts. The initial white blood cell count recorded was 10,700 cells per microliter (reference range 3,500–16,000 cells per microliter). Four more complete blood tests showed white blood cell counts ranging from 1,200 to 1,900 cells per microliter. The initial neutrophil count was 8560 cells per microliter (reference range 2500-8500 cells per microliter). The other four neutrophil counts ranged from 696 to 1292 cells per microliter. The initial lymphocyte count was 1177 cells per microliter (reference range 1200-8000 cells per microliter). The other four lymphocyte counts ranged from 330 to 532 cells per microliter.
3.6. Molnupiravir as primary therapy
A small group of four cats were treated with unlicensed molnupiravir as sole therapy for suspected FIP, as shown in Table 3. Three of them reportedly chose molnupiravir over the unlicensed counterpart GS-441524 due to financial constraints. Cat no. 29 received 12 weeks of oral molnupiravir 12 mg/kg twice daily prior to the treatment shown in Table 3. This cat was disease-free for less than one week prior to restarting oral molnupiravir 19 mg/kg twice daily for 10 weeks.
Cat
Clinical symptoms at the beginning of treatment
Brand name
Initial dosage and frequency
Final dosage and frequency
Duration of treatment (weeks)
Time to improve
Persistent clinical symptoms
Conclusion
Adverse effects
* 27
Hiding, lack of socialization, lethargy, anorexia, URI, vomiting, weight loss
Aura 2801
19 mg/kg twice a day
19 mg/kg twice a day
10
less than 1 week
none
clinical remission
none
28
Anorexia, difficulty walking, distended abdomen, hiding, lack of socialization, lethargy
Aura 2801
8 mg/kg twice a day
8 mg/kg twice a day
13
in two weeks
none
clinical remission
none
29
Anisocoria, blindness, eye color changes, anorexia, hiding, lack of socialization, urinary incontinence, lethargy,
Aura 2801
10 mg/kg twice a day
10 mg/kg twice a day
13
in two weeks
none
clinical remission
none
30
Hiding, lack of socialization, lethargy, pale gums, weight loss
Aura 2801
10 mg/kg twice a day
12 mg/kg twice a day
10
in two weeks
none
clinical remission
none
Table 3. Treatment and outcome characteristics of 4 cats receiving unlicensed molnupiravir as primary therapy.* They received two rounds of molnupiravir treatment; the first round is documented in Table 1.
All four cats were treated with oral molnupiravir Aura 2801 at a mean starting dose of 11.75 mg/kg twice daily (range 8-19 mg/kg) and a mean final dose of 12.25 mg/kg twice daily (range 8-19 mg/kg ). The median duration of treatment was 11.5 weeks (IQR 10-13), with two cats treated for 10 weeks and two cats treated for 13 weeks. A Mann-Whitney test was performed and no significant difference was found between the median duration of molnupivir as rescue therapy (12) and the duration of molnupivir as initial therapy (11.5) (p = 0.692). All owners reported seeing clinical improvement within two weeks and one cat showed improvement within one week. All cats survived the treatment, were disease-free at the time of publication, and no adverse effects of the treatment were reported.
3.7. Molnupiravir by type of FIP
The above information was collected for all 30 cats and then further divided according to the clinical forms of FIP. First, 16 cats with a reported neurological form of FIP were evaluated. Subsequently, the other cats were divided according to ocular (2), effusive (7) and non-effusive (5) forms. The mean starting dose of molnupiravir in the neurological form of FIP was 14.4 mg/kg twice daily, with two cats treated 2-3 times daily. The mean final dosage was 16.4 mg/kg twice daily, with two cats treated 2-3 times daily. The most commonly used initial and final dosage was 12 mg/kg twice daily. Median duration of treatment for neurological FIP was 12 weeks (IQR 10-12,641).
In the two remaining cases of ocular FIP, the mean initial dose was 11 mg/kg twice daily and the mean final dose was 13.5 mg/kg twice daily. The treatment lasted an average of 16.5 weeks. Seven cases of effusive disease were treated with a mean initial dose of 10.5 mg/kg twice daily and a mean final dose of 11.1 mg/kg twice daily. Treatment lasted an average of 13 weeks (IQR 12–16). Five non-effusive cases were treated with a mean initial dose of 10.6 mg/kg twice daily and a mean final dose of 12.8 mg/kg twice daily. One cat was treated once a day. The average duration of treatment was 10 weeks (IQR 8.5-13.5).
3.8. Costs and owner satisfaction
The majority of cats in this study were switched to unlicensed molnupiravir due to treatment failure/relapse or insufficient response. In addition to cats that relapsed or did not respond to unlicensed GS-441524-based therapy, one cat did not tolerate the injectable form of GS, and three owners were restricted due to the cost of treatment. Owners were not required to disclose the financial cost of treatment; this information was provided on a voluntary basis only. In addition, “0” responses reported in the report were not included in the calculation of the following averages because it was not possible to distinguish whether “0” represented no cost or unknown cost. The average reported cost for the first round of GS-441524-based therapy was $3448.83, and similarly, the average reported cost for the second round of GS-441524-based therapy was $3509.09. Only 4 owners reported paying for molnupiravir treatment, while 16 others reported “0” (or no cost/cost unknown). The overall average for the 20 owners who responded to the survey question regarding financial costs (including “0” responses) for molnupiravir was $209. The average cost for the four owners who did not respond with a “0” response was $1045. While 90 % owners reported being “very” or “somewhat” satisfied with their experience of their cat’s molnupiravir treatment, three were “very dissatisfied” with their experience. Unfortunately, no explanation was provided for the reported dissatisfaction.
4. Discussion
In this work, we describe the first known use of unlicensed molnupiravir for the treatment of suspected FIP in cats based on owner-reported data. For the treatment of cats using unlicensed molnupiravir as primary therapy for suspected FIP, the combined data from this study suggests that dosing at 12 mg/kg twice daily for approximately 12 weeks is effective in achieving clinical remission. For the treatment of cats receiving molnupiravir as rescue therapy when failing or relapsing after GS-441524-based therapy, the combined data from this study suggests that dosing at 12-15 mg/kg twice daily for 12-13 weeks is effective in achieving clinical remission. However, when broken down by clinical form of FIP, it was found that neurological cases of FIP were generally treated with a higher dosage than the average for all types of FIP. Ocular, effusive and non-effusive cases were treated with a dosage of around 12 mg/kg twice daily, with some variations. Therefore, dosing of 15 mg/kg molnupiravir twice daily for 12 weeks appears to be effective for neurological cases of FIP. For ocular, effusive, and non-effusive cases, 12 mg/kg molnupiravir twice daily for 12–13 weeks appears to be effective.
These data are somewhat inconsistent with the proposed treatment protocol of the company manufacturing the unlicensed molnupiravir under the trade name HERO Plus 2801. The recommended dosage in the package insert is 25 mg/kg once daily for effusive and non-effusive FIP, 37.5 mg/kg once daily for ocular FIP, and 50 mg/kg once daily for neurological FIP [9]. The package insert for HERO Plus 2801 also reports preliminary results from a study entitled “Effect of Oral Nutrition Therapy on Survival and Quality of Life in Feline Infectious Peritonitis,” which included 286 cats diagnosed with FIP. According to this package insert, 28 cats were cured after 4 weeks of treatment and 258 cats were cured after 8 weeks of treatment, with no deaths at the time of writing [9]. Data from this study have not yet been published in the scientific literature.
However, the cats in this study were using molnupiravir from a different supplier, Aura, which did not provide specific treatment recommendations. The treatment protocols used were therefore based on advice and information shared in groups on social networks, worksheets published on the Internet [10,11] and information on possible adverse effects contained in information published as part of human drug approval applications [12].
The molnupivir treatment protocol derived from this study more closely matches an independently designed protocol [10] published on the Internet. Based on data from in vitro cell cultures of EIDD-1931 and EIDD-2801, laboratory and field studies of GS-441524, and human pharmacokinetic studies, these authors extrapolated the effective dosage of oral molnupiravir [10]. Their calculations suggested a dosage of 4.5 mg/kg every 12 h for effusive and non-effusive FIP, 8 mg/kg every 12 h for ocular FIP, and 10 mg/kg every 12 h for neurological FIP [10]. Although the dosage in this study was generally higher than the dosage suggested by the cited authors, the high survival rate and low relapse rate at the time of the study termination suggest that the manufacturer's unlicensed recommendations may not represent the lowest effective dosage. Ultimately, controlled scientific experiments are greatly needed to evaluate the lowest effective dosage of molnupiravir in cats with suspected FIP.
Several cats were treated with Aura 1931, which is the active metabolite of molnupiravir, EIDD-1931. The reported dosages used were in a similar range to those reported for molnupiravir. Nominally, because the molecular weight of EIDD-1931 is lower than that of EIDD-2801, these cats received more active drug than cats using molnupiravir. However, a previous study showed decreasing oral bioavailability with increasing doses in mice. Therefore, the difference in bioavailability may not be proportional [13]. Pharmacokinetic studies of both molnupiravir and EIDD-1931 in cats are unfortunately unknown.
No adverse effects were reported in the package insert for HERO Plus 2801, which is contrary to what was reported in this study. Among the reported side effects of molnupiravir, the most prominent were drooping ears, hair loss, and severe leukopenia. No skin or follicular lesions have been reported in the human medical literature to match the whisker shedding or ear folding reported here. However, it should be noted that the cats that experienced these side effects received the two highest doses of molnupiravir shown in this study: 23 mg/kg three times daily and 30 mg/kg twice daily.
Severe bone marrow toxicity was reported in dogs during a 28-day study that was discontinued due to severe drug effects [12]. At the dosage of 17 mg/kg/day and 50 mg/kg/day, all hematopoietic cell lines were affected [12]. Cat no. 22 received a maximum dosage of 23 mg/kg three times daily, which was much higher than the toxic dosage in dogs of 17 mg/kg once daily. In the study group with a dose of 17 mg/kg, the possibility of reversibility was noted when the treatment was stopped [12].
There are concerns about the content of unlicensed brands of molnuviravir, as these brands are not currently regulated and often do not list the actual ingredients. The Hero brand (same manufacturer as HERO Plus 2801) shown in Figure 3 was analyzed by our group in December 2021 through Toxicology Associates Inc. (Columbus, OH). It was found to contain 97.3 % of molnupivirus, with no other contaminants detected. The Aura 2801 product used by the majority of participants in this study was analyzed in September 2022 by the same laboratory. It was found to contain 96.8 % of pure molnupivirus. A more controlled assessment of the actual content and purity of the unlicensed preparations of both GS-441524 and molnupiravir is of great interest to the veterinary community and is an active research topic in our group.
Figure 3. Images of Hero brand unlicensed molnupiravir packaging.
Some limitations of this study result from the retrospective nature and legality of the therapies used. First, all data used in this study were obtained based on owner reports. Working closely with the owners and administrators of the social media websites that supported this group enabled a better understanding and interpretation of many of the survey responses. Due to the lack of a definitive ante-mortem diagnosis of FIP available for practical use, it was also not possible to confirm that the cats included in this study had FIP. In addition, the data are likely to be biased toward positive outcomes and may be burdened by recall error. During the distribution phase, a potential study participant responded by requesting to be removed from our email list and stating that he did not wish to participate in the study. Their cat did not respond to molnupiravir treatment and was eventually euthanized. We assume that others may have had the same feeling, since three other potential participants did not respond to the invitation to the study. This may have narrowed the number of participants with an adverse outcome and falsely inflated apparent survival rates. Therefore, the data presented here are intended to serve as evidence of the feasibility of using molnupiravir as primary or rescue treatment for FIP, not as an indication of the true rate of efficacy.
In cats using unlicensed molnupiravir as rescue therapy, the cause of failure to respond or relapse after GS-441524-based therapy was not determined. It could be related to the quality of the drug, the resistance of the virus or another factor. As there is currently no testing or regulation in the US, unlicensed versions of GS-441524 or GC376 may be of insufficient purity or concentration, leading to treatment failure. Another possible cause is natural or acquired resistance to GS-441524. These two causes may also be linked, as acquired resistance may be promoted when an insufficient amount of antiviral is used in treatment, for example with low-quality drugs.
A recent paper found no drug-induced viral mutations of SARS-CoV-2 during molnupivir treatment [14]. This suggests that SARS-CoV-2 is unlikely to develop resistance to molnupiravir. Therefore, treatment with molnupiravir may be similarly unlikely to induce FIPV resistance, making it an attractive therapeutic option.
However, there is clearly a need for (1) a legal (in the United States and elsewhere) alternative to unlicensed treatment with GS-441524 and (2) the availability of alternative rescue drugs, either alone or in combination, after failure of GS-441524 treatment. Molnupiravir has the potential to fill both of these gaps, and this is the first known report of its use in cats in the literature. Nevertheless, unlicensed preparations may continue to be used for the treatment of FIP given the cost and the widely established networks available for their acquisition.
In conclusion, according to owner-reported data, unlicensed molnupiravir appears to be an effective treatment for suspected FIP, both as a first-line and rescue treatment. At a dose of 12–15 mg/kg every twelve hours, minimal side effects are reported and it provides a survival benefit with clinical resolution of FIP. Although the experiences of these owners in treating and possibly curing cats with FIP are unconventional and potentially illegal, they are undeniably remarkable and there is much to learn from the experiments these “citizen scientists” are conducting. In reporting these experiences, we aim to provide a basis for investigating molnupiravir for use in cats with suspected FIP and to document a phenomenon of “group health” that our profession should not ignore.
Supplementary materials
The following supplementary information can be downloaded at https://www.mdpi.com/article/10.3390/pathogens11101209/s1 Supplementary Data S1: retrospective review of molnupiravir trials; additional data S2: abbreviated diary of clinical history cat. no. 6; supplementary data S3: Cat #21 abbreviated clinical history log.
References
Felten, S .; Hartmann, K. Diagnosis of Feline Infectious Peritonitis: A Review of the Current Literature. Viruses2019, 11, 1068. [Google Scholar] [CrossRef] [PubMed]
Pedersen, NC; Kim, Y.; Liu, H.; Kankanamalage, ACG; Eckstrand, C.; Groutas, WC; Bannasch, M.; Meadows, JM; Chang, K.-O. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. J. Feline Med. Surg.2018, 20, 378–392. [Google Scholar] [CrossRef] [PubMed]
Pedersen, NC; Perron, M .; Bannasch, M .; Montgomery, E .; Murakami, E .; Liepnieks, M .; Liu, H. Efficacy and safety of the nucleoside analog GS-441524 for the treatment of cats with naturally occurring feline infectious peritonitis. J. Feline Med. Surg.2019, 21, 271–281. [Google Scholar] [CrossRef] [PubMed]
Jones, S.; Novicoff, W.; Nadeau, J.; Evans, S. Unlicensed GS-441524-Like Antiviral Therapy Can Be Effective for At-Home Treatment of Feline Infectious Peritonitis. Animals2021, 11, 2257. [Google Scholar] [CrossRef] [PubMed]
Merck & Co., Inc. Authorized for Emergency Use in the Treatment of COVID-19. Lagevrio. 2022. Available online: https://www.lagevrio.com/patients/ (accessed on 26 August 2022).
Gordon, CJ; Tchesnokov, EP; Schinazi, RF; Götte, M. Molnupiravir promotes SARS-CoV-2 mutagenesis via the RNA template. J. Biol. Chem.2021, 297, 100770. [Google Scholar] [CrossRef]
Singh, AK; Singh, A.; Singh, R.; Misra, A. Molnupiravir in COVID-19: A systematic review of literature. Diabetes Metab. Syndr. Clin. Res. Rev.2021, 15, 102329. [Google Scholar] [CrossRef] [PubMed]
Khoo, SH; Fitzgerald, R.; Fletcher, T.; Ewings, S.; Jaki, T.; Lyon, R.; Downs, N.; Walker, L.; Tansley-Hancock, O.; Greenhalf, W.; et al. Optimal dose and safety of molnupiravir in patients with early SARS-CoV-2: A Phase I, open-label, dose-escalating, randomized controlled study. J. Antimicrob. Chemother.2021, 76, 3286–3295. [Google Scholar] [CrossRef] [PubMed]
European Medicines Agency. Committee for Medicinal Products for Human Use (CHMP) Assessment Report: Use of Mol-Nupiravir for the Treatment of COVID-19. 2022. Available online: www.ema.europa.eu/contact (accessed on 8 October 2022).
Painter, GR; Bowen, RA; Bluemling, GR; DeBergh, J.; Edpuganti, V.; Gruddanti, PR; Guthrie, DB; Hager, M.; Kuiper, DL; Lockwood, MA; et al. The prophylactic and therapeutic activity of a broadly active ribonucleoside analog in a murine model of intranasal Venezuelan equine encephalitis virus infection. Antivirus. Res.2019, 171, 104597. [Google Scholar] [CrossRef] [PubMed]
Fletcher, T.; Ah Donovan-Banneld, I.; Penrice-Randal, R.; Goldswain, H.; Rzeszutek, A.; Pilgrim, J.; Bullock, K.; Saunders, G.; Northey, J.; Dong, X.; et al. Characterization of SARS-CoV-2 genomic variations in response to mol-nupiravir treatment in the AGILE Phase IIa clinical trial. Res. Sq.2022. [Google Scholar] [CrossRef]
You are all familiar with the standard treatment for FIP in Australia which uses remdesivir (IV or SCI), GS-441524 (tablets) and mefloquine. These three drugs represent the basic equipment for feline veterinarians in Australia, although the specific details and treatment regimens vary from cat to cat according to their clinical signs and the preferences of the attending physician and the financial resources of the owner. Remdesivir has the advantage of being suitable for both intravenous and subcutaneous treatment, which may be useful in some cases of advanced disease or in cases where the abdominal disease is so extensive that there is concern about how much GS-441524 will be absorbed. I suppose no one uses the polyprenyl immunostimulant anymore despite recent work by the Edinburgh group showing some effectiveness.1
Figure 1: Hong Kong British Shorthair cat with moist (effusive) FIP. Photos courtesy of Chris Simpson.
In fact, some doctors and clients prefer to skip remdesivir altogether and go straight to GS-441524 tablets, which are less expensive, forgoing the need for high-cost hospitalization for several days of intensive treatment. The optimal timing of parenteral administration of remdesivir before switching to GS-441524 tablets is debated. We initially recommended two weeks of parenteral treatment, but for example colleagues at the Royal Veterinary College give remdesivir intravenously for 4-5 days and then switch to oral GS-441524.
Mefloquine is a useful drug that can be used in combination with GS-441524, or it can be given when owners can no longer afford the high cost of treatment, at a time when the cat is doing well but is probably not yet cured. There is some debate about the best dosing regimen for this repurposed drug. The original work suggested that ¼ tablet of Larium (62.5 mg) given twice weekly was adequate, while I prefer 20 to 25 mg per cat once daily. I often start this dose in cats near the end of GS-441524 treatment, and I give mefloquine for several months to give the cat’s immune system a little more time to “clear out” any FIP virus that is hiding in the mononuclear phagocytic cells.
Figure 2: High protein fluid from the abdominal cavity of the cat in Figure 1.
I have found that treatment regimens based on these drugs usually result in successful treatment of kittens and cats with FIP, although individual cases can be very challenging.
In my opinion, the biggest obstacle to successful treatment is the very high cost of treatment. Another problem is the requirement of 84 days of treatment to completely eliminate the virus.
The drugs themselves are very expensive, especially in adult cats or patients with CNS disease (which require higher mg/kg doses for drugs to penetrate the CNS), and this includes not only the very high cost of the drugs, but also the cost of initial stabilization and ongoing consultations for monitoring. As a result, treatment is financially unaffordable for many owners, often from the outset, and if resistance to the FIP virus develops during treatment, the requirement for very high doses, often over a long period of time, makes treatment difficult for even the most dedicated owner.
The owners tried to circumvent the high cost of treatment by using the black market drug GS-441524, which is commonly available from many suppliers. Although it is not legal, owners and especially cat breeders have obtained it in large quantities and many cats have been saved by these drugs.2 The problem is that we are not sure of the actual doses or the quality or the product that is given in different colored tablets, and our testing has shown that the dose, even if indicated on the package, may be higher or lower than the value indicated by the manufacturer. In addition, we cannot assess the differences between individual batches of black market drugs. Therefore, most Australian vets advise clients to use legal products provided by BOVA Australia and the supply chain is reliable and regular quality control ensures that each tablet contains 50mg of GS-4415624 as stated.
Figure 3: X-ray of the cat from Figure 1 with wet FIP.
The COVID pandemic has led to tremendous research into the prevention and treatment of coronavirus disease, and two oral products for the oral treatment of human patients infected with SARS-CoV2, namely molnupiravir and paxlovid, are now commonly available in Australia and elsewhere.3 Niels Pedersen provided the SOC FIP on his website a summary of the history of molnupivirus that I have attached to this monograph. The key part is cut and pasted below (with some editing):
As expected, molnupiravir was recently tested in cats with FIP by at least one Chinese vendor of GS-441524, and preliminary results were reported on the FIP Warriors CZ/SK website. The field study consisted of 286 cats with various forms of naturally occurring FIP seen at companion animal clinics in the US, UK, Italy, Germany, France, Japan, Romania, Turkey and China. There were no deaths among the 286 cats that participated in the tests, including seven cats with ocular (n=2) and neurological (n=5) FIP. Twenty-eight of these cats were cured after 4-6 weeks of treatment and 258 after 8 weeks. All treated cats remained healthy after 3-5 months, which is the period during which cats that were not successfully treated would be expected to relapse.
These data provide compelling evidence for the safety and efficacy of molnupiravir for cats with various forms of FIP. However, we hope that this field study will be written up in manuscript form, submitted for peer review and published. Nevertheless, it is now marketed to owners of cats with FIP. At least one other major marketer of GS-441524 is also interested in using molnupiravir to treat FIP, indicating demand for additional antiviral treatment in cats with FIP.
The safe and effective dosing of molnupiravir in cats with FIP has not been published. However, at least one vendor from China provided some pharmacokinetic and field test data for molnuparivir in cats with naturally occurring FIP in their promotional leaflet for a product called Hero-2801. In 28/286 cases, they received this drug at a dose of 30-40 mg/kg every 24 hours, ie the equivalent of 15-20 mg/kg every 12 hours. For comparison, the dose for humans is 800 mg every 12 hours or about 10 mg/kg per day.
Figure 4: Data provided on “Hero 2018” by FIP Warriors CZ/SK – EIDD-2801 (Molnupiravir). https://www.fipwarriors.eu/en/eidd-2801-molnupiravir/
Dosage recommendations seem to vary. It was originally proposed:
FIP: 25mg/kg q24h
Ocular FIP: 37.5 mg/kg q24h
Neurological FIP: 50 mg/kg q24h
The duration of treatment is 5-10 weeks, depending on the severity of the disease and the specific cat.
Later, this proposal was modified based on input from Niels Pedersen and the UC Davis group:
FIP: approximately 5-7 mg/kg q12h for 84 days.
Ocular FIP: 8-10 mg/kg q12h for 84 days.
Neurological FIP: 10-15 mg/kg q12h for 84 days.
These recommendations are based on assumptions based on published information and more field experience with molnupiravir is needed. Sam Evans has just presented some data on the use of molnupiravir for salvage therapy at the ISCAID conference in Glasgow. An active clinical trial involving Brian Murphy and Krystal Regan is underway at the University of California, Davis, to determine the optimal dosage and dosing interval for molnupiravir, beginning in July 2022.
It is questionable whether molnupiravir will prove safer and more effective than GS-441524 in the treatment of FIP, but a third antiviral could prove extremely useful in preventing resistance to GS-441524 (as a cocktail of antivirals with different resistance profiles) or in treating cats that they no longer respond well to GS-441524. The big unknown is whether molnupiravir will be free of long-term side effects.
As the dosages given in the trials seem to be somewhat questionable, I used 10mg/kg twice daily, but note that test cats are claimed to have received 100mg/kg once daily with no detectable adverse effects (see Figure 4) .
Key fact: I have been using molnupiravir in selected patients for about 2 months at a dose of 10 mg/kg twice a day. It may be prudent to increase this dose to 15 mg/kg twice daily, particularly in CNS disease. Higher doses seem likely to be safe and possibly more effective, but I'm reluctant to recommend them until we get more long-term data.
Finally, and really on a side note, paxlovid is a combination of two drugs given at the same time, with one used to inhibit the metabolism of the other; I can find no precedent for its use in cats, which is a shame because in humans it is the more effective of the two oral drugs available for COVID. It costs about the same in Australia as molnupiravir, but is comparably more expensive than molnupiravir when purchased from websites in India. It could prove to be a very useful drug for treating FIP if it is proven safe. In humans, one of the problematic side effects is a foul taste in the mouth, so-called “paxlovid mouth”, which could prove disastrous if it were to occur in cats, as they tend to foam at the mouth.
So what is the place of molnupiravir in the therapy of kittens and cats with FIP? How to get it? How much does it cost?
Anyone can buy molnupiravir for personal use or for use in cats by obtaining a prescription and presenting it at a pharmacy. The price is approximately $1,146.39 (https://www.pbs.gov.au/medicine/item/12910L) subject to pharmacy surcharge. The trade name is Lagevrio (Merck Sharp & Dohme) and the box contains 40×200 mg capsules. The drug was provisionally approved by the TGA in February 2022 for the treatment of COVID-19 in adults who do not require oxygen and are at risk of progression to severe COVID-19.
To treat a 4 kg cat at 10-15 mg/kg bid, you need 80 mg to 120 mg daily for 84 days or 6,720 to 10,080 mg. A box of 40x200 mg capsules is 8,000 mg, so if you factor in the preparation fee for a suitable dose for a cat - the high dose treatment costs about $1100-$2200, which is quite a bit cheaper than GS-441524 or remdesivir. So now we have an alternative treatment to the one we are currently using. What is the evidence? Are pharmacokinetics well known in cats? The answer to both is - we don't know for sure because the evidence hasn't been peer-reviewed, but compelling unpublished anecdotal information suggests it's an effective therapy. And we'll probably have good PK info from the Davis group within a year.
And now it gets a little more complicated! Australia has a system that allows people to import medicines from overseas for their own personal use and that of their family members. The reason is that human patients who are prescribed expensive unregistered medicines, and therefore medicines not covered by the PBS, have to find their own way of getting these medicines. It is not known how many Australians import medicines, but it is legal under the personal import scheme. Perhaps the best example is the “FixHepC Buyers Club” (https://fixhepc.com/), founded by Australian infectious disease doctors and general practitioners Drs John and James Freeman.5 Before hepatitis C drugs were subsidized by the PBS, thousands of Australians used this buyers' club to import affordable hepatitis C drugs at 1-2 % of the retail price. The initiative was supported by the Australian Society for HIV, Viral Hepatitis and Sexual Health Medicine, and many Australian doctors directed patients to buy their medication this way.
"They each paid $1,000 to $2,000 to get better, instead of the $84,000 that Gilead Sciences charged in America. It's still expensive, but at least it's affordable for many."
However, it comes with some risk. The online market for medicines is poorly regulated, operating across jurisdictions and substandard products are common. Some estimates suggest that up to 25% of medicines in circulation outside high-income countries are substandard. The biggest risk is insufficient quantities of the active substance, which can lead to unintentional undertreatment. This is a difficult area for doctors to navigate, both clinically and medico-legally. It is unclear how far a doctor’s duty of care extends. The current code of conduct states that good medical practice includes “upholding the patient’s right to access the necessary level of healthcare and, wherever possible, assisting them to do so.” There is no reason to believe that this would not also include helping patients to import medicines they would otherwise be unable to afford, if it is clearly in their best interest to do so.
My own opinion is that this is the same situation for small animal vets involved in the treatment of cats with FIP, and the vet oath taken by recent graduates would support this claim.
So how can a client order molnupiravir to treat their cat with FIP?
Figure 5; Screenshot of correspondence with the manufacturers of Molcovir 200 mg
Go to the internet and find the URL of the website called IndiaMART https://www.indiamart.com/ and search for Molcovir 200mg - currently the URL is: https://m.indiamart.com/isearch.php?s=Molcovir +200mg&prdsrc=1&countryiso=AU&qu-cx=1&stype=attr=1
They will take your information and ask what medicine you are interested in. They will then send your inquiry to several pharmacies and ask you for quotes. The manufacturers we have the most experience with are Dolphin pharmaceuticals and Mediseller, one of which accepts payment via PayPal instead of using your credit card. They will advise you that customs is your own business, but they will do their best to help you label the medication so that it is clear that it is not an illegal drug. You can't buy anything until the quote comes back to you. Figure 5 shows a screenshot of the correspondence with the company. Once you accept the quote, they will then arrange payment and then shipping. Currently, a five-pack purchase costs US$30 (AU$42) per pack plus US$65 shipping. Treating a 4kg cat with FIP CNS at 10-15mg/kg bid for 84 days is likely to be in the range of AUD$100-150. The cost of treating the same cat with remdesivir/GS-441524 is $6,550 when drug costs alone are considered.
So the cost of treating a cat with molnupiravir from India under the personal importation scheme will be about 2 % of the cost of conventional GS-441524 treatment. This means that no cat has to be euthanized due to the high cost of treatment –
Figure 6; Generic molnupiravir (Molcovir 200 mg capsules) purchased from IndiaMART and imported to Australia after customs clearance
The practical question is how to obtain molnupiravir in time. When purchased from this website, it takes approximately 3 weeks for the drug to reach Australia and clear customs. One option would therefore be to start treatment with remdesivir and/or GS-441524 and switch to molnupiravir as soon as the drug arrives. An alternative is to set up a “buyers’ club”, perhaps with the support of the Australian and New Zealand College of Veterinary Science Pharmacology chapter, to be the source of molnupiravir until the client can arrange to import their own supplies. Another option would be for a compounding pharmacy to import the drug from a reliable manufacturer and adapt it to a size suitable for cats and kittens, perhaps as a 60 mg metered tablet that can be split in half.
What is the best way to treat a cat with FIP in 2022 in Australia?
This question is not as simple as it might seem. Remdesivir and GS-441524 have had much success as a treatment for FIP in kittens and cats, and we are also comfortable with low-dose mefloquine as an add-on to consolidate treatment. However, most of us who have assisted in the treatment of cats know of cases where the FIP virus has developed acquired mutational resistance during treatment. This can often be worked around by increasing the doses of GS0-441524 administered, but many owners simply cannot afford this procedure.
Thus, the routine use of combination therapy of FIP cases using GS-441524 and molnupiravir can be recommended, and it is indeed possible that the combination therapy could be more effective and faster because it attacks two different targets in the FIP virus, and this could potentially lead to a shorter duration of treatment. The idea of 84-day therapy comes from the seminal work of Neil Pedersen and perhaps the assumption that the lifespan of macrophages in tissues is about 84 days, so to completely eliminate all the intracellular virus that is hiding in the mononuclear phagocyte system, we need to treat for longer than 84 days of lifespan macrophage.
However, for many clients, the huge cost of treatment is an insurmountable problem, and for many, and I dare say most clients, the ability to treat their kitten or cat for $200 (drug costs) will be the most important argument.
What is the disadvantage of molnupivir? The big unknown is whether molnupiravir will not have long-term toxic effects, since the active substance, N4-hydroxycytidine, is an extremely potent mutagen and the treatment time for FIP is much longer than the 5 days recommended for the treatment of Covid-19 in human patients. The probability of side effects is therefore theoretically higher. To me, that's a completely theoretical risk, but it's something we have to monitor in the patients we treat, being aware of the possibility of later cancer in some cases.
References
1. Černá, P.; Ayoob, A.; Baylor, C.; Champagne, E.; Hazanow, S.; Heidel, RE; Wirth, K.; Legendre, AM; Gunn-Moore, DA Retrospective Survival Analysis of Cats with Feline Infectious Peritonitis Treated with Polyprenyl Immunostimulant That Survived over 365 Days. Pathogens2022, 11, 881. https://doi.org/10.3390/pathogens11080881
2. Jones, S.; Novicoff, W.; Nadeau, J.; Evans, S. Unlicensed GS-441524-Like Antiviral Therapy Can Be Effective for At-Home Treatment of Feline Infectious Peritonitis. Animals2021, 11,2257. https://doi.org/10.3390/ani11082257
3. Atmar RL, Finch N. New Perspectives on Antimicrobial Agents: Molnupiravir and Nirmatrelvir/Ritonavir for Treatment of COVID-19. Antimicrob Agents Chemother. 2022 Aug 16;66(8):e0240421. doi: 10.1128/aac.02404-21. Epub 2022 Jul 12. PMID: 35862759; PMCID: PMC9380556.
4. https://ccah.vetmed.ucdavis.edu › files › inline-files (this is a fantastic review of this subject, although some of the arithmetic is wrong. Highly recommended for people who are interested in drug development and action.)
Arjun N. Sweet, Nicole M. André, Alison E. Stout, Beth N. Licitra and Gary R. Whittaker Julia A. Beatty, Academic Editor and Séverine Tasker, Academic Editor
The emergence of severe acute respiratory syndrome 2 (SARS-CoV-2) has led the medical and scientific community to address questions regarding the pathogenesis and clinical presentation of COVID-19; however, relevant clinical models other than humans are still lacking. In cats, the ubiquitous coronavirus, described as feline coronavirus (FCoV), can manifest as feline infectious peritonitis (FIP), a leading cause of mortality in young cats characterized by severe systemic inflammation. The diverse extrapulmonary symptoms of FIP and the rapidly progressive course of the disease, together with the proximate etiologic agent, represent a degree of overlap with COVID-19. This article reviews the molecular and clinical relationships between FIP and COVID-19. Although there are key differences between the two syndromes, these similarities encourage further investigation of feline coronaviruses as a naturally occurring clinical model for human coronavirus disease.
In the 1960s, feline infectious peritonitis (FIP) was described as a disease in domestic cats and was subsequently found to be of viral etiology, specifically feline coronavirus (FCoV). [1,2]. In most cats, infection with FCoV results in mild to inconspicuous clinical signs, but a small proportion of cats develop severe disease and succumb to the systemic form of the disease known as FIP [3]. In the years since the discovery of FCoV, many features of FCoV have remained misunderstood. Similarly, the COVID-19 pandemic, caused by the emergence of SARS-CoV-2, has raised many equally challenging questions regarding pathogenesis, transmissibility, and treatment. The widespread transmission of FCoV/SARS-CoV-2 and the insidious onset of severe symptoms in both FIP and COVID-19 limit the ability to detect the disease early—what may begin as mild or even mild clinical signs or symptoms can quickly lead to systemic disease [3,4]. We believe that FIP may represent a valuable, naturally occurring extrapulmonary model of COVID-19.
Both FCoV and SARS-CoV-2 belong to the family Coronaviridae [4,5], although to different genera (Figure 1). FCoV, together with similar animal coronaviruses such as canine coronavirus (CCoV) and porcine gastroenteritis virus (TGEV), belong to the genus alpha-coronaviruses. Community respiratory (CAR) human coronaviruses 229E and NL63 are also included in the genus Alphacoronaviruses. [6], the latter being associated with the common cold, hail and possibly Kawasaki disease in children [7]. In contrast, SARS-CoV-2, together with SARS-CoV (the causative agent of the severe acute respiratory syndrome outbreak in 2002-2003) and the Middle East respiratory syndrome coronavirus (MERS-CoV) belong to the genus betacoronaviruses [8], wherein SARS-CoV-2 and SARS-CoV belong to line B (sarbecovirus) and MERS-CoV belong to line C (merbecovirus). Less related beta coronaviruses include human coronavirus CAR OC43 (associated with the common cold), mouse hepatitis virus (MHV), and bovine coronavirus, which is associated with pneumonia and diarrhea in cattle; these viruses are in line A (embekovirus).
Figure 1 Phylogenetic tree of spike proteins of selected coronaviruses. The phylogenetic tree of maximum likelihood was constructed using the MEGAX program (100 bootstraps) from multiple alignment of spike protein sequences. Tip amino acid sequences were obtained from GenBank NCBI. Relevant numbers are: transmissible gastroenteritis virus / TGEV (P07946), severe acute respiratory syndrome coronavirus 2 / SARS-CoV-2 (YP_009724390.1), middle eastern respiratory syndrome / MERS-CoV coronavirus (AFS88 /36) 1. -1 (ACN89742), severe acute respiratory syndrome coronavirus / SARS-CoV (AAT74874.1), feline coronavirus / FCoV-Black (EU186072.1), bovine coronavirus / BCoV (P15777), canine coronavirus / CCo37.14) , human coronavirus / HCoV-OC43 (NC_006213.1), HCoV-229E (NC_002645.1) and HCoV-229E (NC_002645.1).
FCoV can be classified in two ways, the first of which relates to the form of the disease. Feline enteric coronavirus (FECV) is thought to cause a mild gastrointestinal form of the disease, while feline infectious peritonitis virus (FIPV) is associated with a fatal systemic infection known as FIP. [3]. FIPV differs from FECV in its ability to infect and replicate efficiently in monocytes and macrophages [9], causing systemic inflammation. FIPV is associated with a spectrum of clinical outcomes. At one end of the spectrum is effusive or “wet” FIP, which progresses rapidly and involves the accumulation of a high-protein exudate in the abdominal and/or thoracic cavities. At the other end of the spectrum is noneffusive or “dry” FIP, which can affect many organ systems but is usually characterized by neurological and ocular symptoms. Noneffusive FIP generally has a more protracted course of disease and is less common than its effusive counterpart. FCoV can also be divided into two serotypes—type I or type II—based on major differences in the virus’s spike protein, which affect receptor binding and antibody response. [10]. The receptor for FCoV type II is feline aminopeptidase N (fAPN) [11], while the receptor for type I viruses is not identified. Type I FCoV accounts for the vast majority of FIP cases [12].
The classification of SARS-CoV-2 virus into different variants based on genetic mutations is still ongoing as the virus continues to evolve. Viral lines that show the potential for increased transmissibility, treatment resistance, vaccine resistance, or increased morbidity and mortality have been identified as VOCs. The spectrum of diseases associated with COVID-19 is broad, ranging from asymptomatic and mild infections to acute respiratory distress syndrome (ARDS), systemic inflammatory response syndrome (SIRS), and multiorgan failure and death. Systemic inflammation in SARS-CoV-2 is not associated with macrophages and monocytes (as in FIP), but is responsible for a wide range of extrapulmonary symptoms. The SARS-CoV-2 receptor, angiotensin converting enzyme-2 (ACE-2), which plays an important role in the renin-angiotensin system and the development of pro-inflammatory status, appears to be involved. [13]Multisystem inflammatory syndrome (MIS) in children and adults, as well as post-acute course of SARS-CoV-2 infection (PASC), also known as “long COVID”, are potential consequences of COVID-19 infection.
2. Transfer
As a group, coronaviruses are known for their ability to cause both respiratory and intestinal diseases and are usually transmitted in one or both ways. While FCoV is considered faecal-oral and SARS-CoV-2 is primarily respiratory, patients with COVID-19 may excrete the infectious virus in the faeces. [14], often for a long time, and FCoV can easily become infected by the oronasal route, which is a common method of experimentally vaccinating cats [15].
In most cases, FCoV infection is self-limiting, and although the virus can be detected systemically, replication outside the intestinal epithelium is weak. This form of the virus, referred to as FECV, is easily transmitted by the fecal-oral / oronasal route, with common anchors and swallowing of virus particles during purification being common sources of infection. The current understanding of FIP development involves internal mutation: in a small subset of FECV cases, a complex combination of host and viral factors leads to mutation (s) that allow efficient replication in macrophages and monocytes. [16]. These lethal variants are classified as FIPV and are associated with systemic inflammation, organ failure, and death. FIPV is generally considered non-transmissible because factors that increase its tropism on troprophages appear to limit its faecal-oral spread. [17]. FIP outbreaks were recorded in kennels and shelters. In these situations, congestion stress and high levels of virus in the environment may promote the conversion of FECV to FIPV. There is evidence that some FCoV strains may be more susceptible to this rebirth than others [18,19].
SARS-CoV-2 virus infection is primarily targeted at the respiratory epithelium, but as with FCoV, the virus may appear systemically without appropriate symptoms of infection. [20,21]. Asymptomatic individuals are a well-documented source of SARS-CoV-2 [22,23,24] and delivery includes inhalation of aerosols as well as contact with droplets [25]. The incubation times of SARS-CoV-2 and FECV range from 2 to 14 days [26]. The incubation time of FIP is highly variable, influenced by the time to internal mutation and the individual's immune response. The onset of FIP can occur several weeks to months after the initial infection [27,28,29,30]. Multisystem inflammatory syndrome in children (MIS-C), a severe manifestation of SARS-CoV-2, is also delayed by an initial infection with a median onset of 4 weeks. No viral factors have been associated with the development of MIS-C, but an immune-mediated component is thought.
Vertical transmission of FIP via the placenta or milk is considered rare. In the first experimental study in which a suckling cat was infected, one in four kittens succumbed to FIP [28]. Maternal antibodies appear to be effective in preventing transmission until approximately six weeks of age, when antibody levels decrease and kittens are susceptible to fecal-oral transmission. [31]. However, this maternally acquired immunity can be overcome early in life by high levels of exposure to FCoV – a Swiss study showed that kittens in large herds show infection as early as two weeks of age [32,33]. Vertical transmission poses a risk of SARS-CoV-2 infection. Placental transmission is rare but has been documented in fetuses of SARS-CoV-2 infected mothers. [34,35,36], as evidenced by virus detection in amniotic fluid, neonatal blood, umbilical cord blood and placental tissue. Transmission cases have been documented in both early and late pregnancies, but infection of the neonate with SARS-CoV-2 may not always occur in the uterus. Infection can also occur during childbirth or in close contact with the mother. The neonatal outcomes of COVID-19 infected mothers remain under study, and it is difficult to distinguish between the effects of SARS-CoV-2 infection and maternal comorbidities. Nevertheless, neonatal infection does not appear to be without sequelae, with one analysis showing that approximately 50 % infected neonates exhibited COVID-19-related clinical symptoms, including fever and respiratory and gastrointestinal symptoms. [37].
3. General clinical presentation
Clinical signs associated with both FIP and COVID-19 include fever, diarrhea, depression, weakness, anorexia, and dyspnoea. [1]. Typical manifestations of COVID-19 commonly include non-specific symptoms including fever, dry cough, fatigue, dyspnea, and myalgia [38]. Anosmia (loss of smell) and ageusia (loss of appetite) have also been frequently reported in COVID-19 and are more specific symptomatic indicators of the disease. [39]. Pneumonia, acute respiratory distress syndrome (ARDS) and sepsis may occur. Men appear to be at higher risk of developing more severe manifestations of COVID-19 [40,41], with several small studies confirming the same relationship between males and the development of FIP in cats [42,43].
The classic manifestation of FIP is the formation of effusion in the abdomen and / or thoracic cavity; although this manifestation has also been reported in COVID-19 [44], is very rare. In addition, FIP is manifested in various bodily systems that are similar to the extrapulmonary manifestations of COVID-19 (Figure 2 and Figure 3). The most similar feature of both diseases is endothelial dysfunction. Vasculitis is a hallmark of FIP pathology [45,46] with lesions characterized by perivascular edema and infiltration, vascular wall degeneration and endothelial proliferation [47]. In the case of COVID-19, extrapulmonary symptoms are thought to be caused by virus-mediated endothelitis, which leads to vasculitis, especially in veins with minor arterioles [48,49]. In the following sections, we will describe these extrapulmonary symptoms and point out key similarities and differences.
Figure 2 Summary of systemic clinical signs and pathological conditions associated with FIP. FIP is known to be a systemic infection with a variety of manifestations. Summarized possible systemic clinical signs associated with FIP are summarized, which include organ systems that are also affected by COVID-19. The most common symptoms of FIP are highlighted in red.
Figure 3 Summary of systemic clinical signs, symptoms, and pathologies associated with COVID-19. Respiratory symptoms of COVID-19 are the main manifestation of the disease. However, SARS-CoV-2 infection in humans can also result in a variety of extrapulmonary symptoms. Summarized are the systemic clinical signs and symptoms associated with COVID-19, which include organ systems that are also affected by FIP. The most common symptoms of COVID-19 are highlighted in red. ARDS stands for Acute Respiratory Distress Syndrome.
4. Biomarkers
Inflammatory biomarkers are important as prognostic markers in COVID-19 and as a means of differentiating FIP from other diseases. In FIP, IL-6 expression appears to be increased in the ascitic fluid of FIP-infected cats, presumably through increased expression in the heart and liver. [50,51]. Other acute phase proteins are also elevated in FIP infection. Alpha-1-acid glycoprotein (AGP) has been studied as a diagnostic marker of FIP, but may be elevated in other conditions, thereby limiting its specificity. [52,53]. Serum amyloid A (SAA) is another acute phase protein that appears to distinguish between FIPV and FECV infection, with FIPV-infected cats showing higher levels of SAA compared to FECV-infected cats and control cats without SPF. [54], but has limited utility in distinguishing FIP from other effusive conditions [55].
As with FCoV, subjects with severe COVID-19 have higher SAA levels compared to subjects with milder COVID-19. [56]. Higher SAA levels are also reported in patients who died of COVID-19 compared with those who survived [57]. C-reactive protein (CRP) is another marker that has been shown to be a promising biomarker in both FCoV and SARS-CoV-2 infections. Liver CRP synthesis is induced by IL-6 expression in response to inflammation [58] and is increased in FIP cases [59]. Elevated CRP levels in the early stages of COVID-19 are associated with a more severe course of the disease and higher mortality [60,61,62], which led to the recommendation to use it as a prognostic indicator in risk assessment in patients hospitalized for COVID-19. In contrast, one meta-survey found that IL-6 levels were elevated, but at least one order of magnitude lower in patients with COVID-19 than in patients with ARDS and non-COVID-19-related sepsis, suggesting a different mechanism of immune dysregulation. [63].
The D-dimer, although not specific for COVID-19 or FIP, is another interesting biomarker. D-dimer is released upon fibrin breakdown and is used as a clinical tool to eliminate thromboembolism [64]. Thrombotic events have often been documented in COVID-19 in several organ systems [65,66] and elevated D-dimer levels are associated with higher morbidity and mortality [67,68]. Similarly, thrombotic events can occur in FIP, and high levels of D-dimers, along with other symptoms of disseminated intravascular coagulation (DIC), can be observed in the final stages of FIP in both natural and experimental infections. [69,70].
5. Pathophysiology
5.1. Neurological
FIP is one of the major infectious neurological diseases in cats and the symptoms associated with central nervous system (CNS) infection are well documented. [71]. CNS symptoms are reported in approximately 40 % cases of dry FIP and may manifest as nystagmus, torticollis, ataxia, paralysis, altered behavior, altered mentoring, and seizures [72]. The wide range of symptoms supports the conclusion that the infection is not limited to a specific part of the CNS [73]. CNS infection is restricted to the monocyte and macrophage lines and leads to pyogranulomatous and lymphoplasmacytic inflammation, which usually affects leptomening, choroidal plexus and periventricular parenchyma. [74].
Documentation of neurological symptoms associated with SARS-CoV-2 CNS infection is limited compared to other coronaviruses [75]. The symptoms observed range from headache and confusion to seizures and acute cerebrovascular events. [76]. Virus detection in the brain is rare, suggesting that symptoms may not be directly related to CNS infection. Viral particles have been observed in neural capillary endothelial cells and a subset of cranial nerves, although such detection does not correlate with the severity of neurological symptoms. [77]. There is often no clear evidence of direct infection. Instead, inflammatory mediators, such as activated microglia, have been reported to contribute to microvascular damage and disease. [78,79].
Further comparison of the neuroinflammatory properties of SARS-CoV-2 and FCoV may provide new insights into the neurological manifestations of COVID-19. Further understanding of the neurological symptoms associated with SARS-CoV-2 is necessary to understand the progression of COVID-19 and the extent of CNS infection.
5.2. Ophthalmological
Ocular manifestations of FIP are more common in the dry form of the disease [80]. Mydriasis, iritis, retinal detachment, conjunctivitis, hyphema and keratic precipitates have been observed [81]. The most common ocular manifestation of FIP is uveitis, which can affect both the anterior and posterior uvea [80]. Viral antigen can also be detected in epithelial cells of the nitrating membrane, but viral antigen detection does not distinguish between FECV and FIPV [82].
Ocular manifestations of COVID-19 include conjunctivitis, chemosis, epiphora, conjunctival hyperaemia, and increased tear production [83]. Uveitis—a common ocular presentation of FIP—has also been observed in SARS-CoV-2 infection [84,85]. Tear fluid virus detection has led to concerns about ocular transmission in the first months of the COVID-19 pandemic [83,86]. SARS-CoV-2 RNA was detected in tear secretions and was isolated from ocular secretions, supporting the possibility of ophthalmic transmission [87,88]. Interestingly, in the above case study in China, out of 12 patients with ophthalmic symptoms, only 2 patients returned positive conjunctival tests, suggesting limited sensitivity in detecting virus from conjunctival specimens. [83].
5.3. Cardiovascular
Pericardial effusion is a less common manifestation of FIP, but is well documented in the literature [26,89,90,91]. FCoV was detected in the pericardium of cats with recurrent pericardial effusion, which later developed neurological symptoms [92]. Direct FCoV infection of the heart was documented in a 2019 case study of FIP-associated myocarditis with severe left ventricular hypertrophy and atrial enlargement. [93]. Immunohistochemistry (IHC) revealed the presence of FCoV-infected macrophages and associated pyogranulomatous lesions. [26]. Interestingly, severe SARS-CoV-2 infection with evidence of viral replication in the heart and lungs has recently been documented in cats with pre-existing hypertrophic cardiomyopathy (HCM). [94].
Unlike FIP, heart damage associated with SARS-CoV-2 infection appears to be much more prevalent. A study of 187 patients found that 27.8 % cases of COVID-19 showed evidence of myocardial damage, as evidenced by elevated cardiac troponin (TnT) levels. [95]. High levels of TnT were in turn associated with higher mortality. In a retrospective multicenter study of 68 patients with COVID-19, 27 deaths were attributable to myocardial damage and / or circulatory failure as one of the leading causes of mortality, with elevated C-reactive protein and IL-6 levels associated with higher mortality [96]. The increase in such inflammatory biomarkers in the blood suggests that the rapid inflammatory nature of COVID-19 may have a particularly detrimental effect on heart function. Diffuse edema as well as increased wall thickness and hypokinesis have been reported with COVID-19 infection. [97]. Cardiac tamponade was also observed in patients with COVID-19, with SARS-CoV-2 levels detectable in pericardial fluid. [98]. In contrast to FIP, in which direct invasion of FCoV-infected macrophages into the myocardium was observed in myocarditis, myocardial infection with SARS-CoV-2 virus is not clearly associated with mononuclear cell infiltration or myocarditis. [99]. This leads to considerations of multiple systemic factors in adverse cardiac outcomes – particularly dysregulation of inflammatory cytokines. The impact of SARS-CoV-2 infection on the cardiovascular system is an important element in our increasing understanding of the morbidity and mortality associated with COVID-19.
5.4. Gastroenterological
FCoV is excreted in feces and transmitted by the oronasal route. Initial FCoV infection targets the intestinal tract – infection may be subclinical or cats may develop diarrhea and, less commonly, vomiting. The primary infection lasts for several months and the virus can be shed for months to years [100,101]. Colonic epithelial cells appear to serve as a reservoir for persistent infection and excretion. [21]. The symptoms tend to be mild and spontaneous and only a small proportion of the animals go into the FIP stage. Fibrinous serositis and pyogranulomatous lesions with vasculitis are classic FIP lesions and can be found in the small and large intestines of affected cats. [102]. FIP can cause solitary mass lesions in the intestinal wall, although this is considered a rare presentation (26/156 cats in one study) [103]. These are usually found in the colon or ileocecal junction and have a pyogranulomatous character.
Gastroenterological symptoms are often reported with COVID-19 infection. ACE2, the cellular receptor for SARS-CoV-2, is widely expressed in glandular cells of the gastric, duodenal and rectal epithelium. Viral RNA and nucleocapsid were detected in these tissues [104], which supports their suitability for SARS-CoV-2 replication. Gastrointestinal (GI) symptoms range from general anorexia to diarrhea, nausea, vomiting and abdominal pain [105,106]. Excluding a less specific anorexia symptom, multiple meta-analyzes estimate the prevalence of GI symptoms in patients with COVID-19 at approximately 10 % to 20 %, with diarrhea being the most commonly reported symptom. [106,107,108]. Interestingly, GI symptoms in COVID-19 were observed without accompanying respiratory symptoms [105].
Faecal virus excretion is a major concern for COVID-19, as SARS-CoV-2 RNA may continue to be present in faeces even after reaching undetectable levels in upper airway samples. [109]. Although the detection of viral RNA in faeces alone does not necessarily indicate the presence of infectious virions, viable viral particles have been detected in faeces. [110]. The viral antigen persists in the cells of the gastrointestinal tract as well as in the convalescent phase, up to 6 months after healing [20]In one case study, persistent colon infection was associated with persistent gastrointestinal symptoms in a case of “long COVID.” [111], which draws a parallel to the role of the colon epithelium as a reservoir for FCoV.
5.5. Dermatological
Dermatological changes have been reported with both SARS-CoV-2 and FIPV infections. Although papular skin lesions are rare, they are the primary dermatological manifestation of FIP, with several available case reports documenting papules. [81,112,113,114]. On histological examination, pyogranulomatous dermatitis, phlebitis, periflebitis, vasculitis and necrosis were reported in several FIP cases. [81,112,113,114,115].
The first report of dermatological manifestations associated with COVID-19 was recorded at Lecco Hospital in Lombardy, Italy [116]. In this study, 18/88 patients (20.4 %) developed a skin disorder, with 8/18 patients observed at the onset of the disease and 10/18 after hospitalization [116]. Clinical signs included erythematous rash (14/18 patients), diffuse urticaria (3/18 patients) and smallpox-like vesicles (1/18 patients) [116]. Lesions were observed mainly on the trunk (torsion) and pruritus was mild or absent [116]. The continuation of the pandemic brought better characteristics of the first observed dermatological symptoms, as well as the identification of rarer presentations. The most common dermatological manifestation of COVID-19 appears to be rash, often characterized by maculopapular lesions. [117,118]. Another predominant dermatological symptom also appears to be urticaria [118,119]. Importantly, neither rash nor urticaria is specific for COVID-19, which limits their positive predictive value. Varicella-like rash has been observed with SARS-CoV-2 infection and may be more specific due to its low prevalence in viral diseases. In particular, with missing lesions in the oral cavity and pruritus observed in COVID-19-associated rash, together with a previous history of varicella infection, the specificity of this presentation is reinforced. [118].
5.6. Teriogenological
Orchiditis and periorchitis with fibrinopurulent or granulomatous infiltrates as well as hypoplastic testes have been observed in several cases of FIP. [1,26,120]. Inflammatory mediators from the tuna surrounding the testicles caused enlargement of the testicles in cats with FIP [26,120]. In effusive FIP, enlargement of the spinal cord due to edema and peritonitis of tuna was observed [16]. Despite the apparent pathology of the male reproductive system in cats, FCoV has not been detected in sperm, which reduces the likelihood of sexual transmission. [121]. The pathology of the female reproductive system in FIP is less documented in the literature, but macroscopic lesions present in the ovaries of FIPV-infected cats have been observed. The surrounding uterine and ovarian vessels of these cats were surrounded by lymphocytes, macrophages, plasma cells and neutrophils. [122].
As with FIP, the pathology of COVID-19 is manifested in the male reproductive system. One study examining the testes of 12 COVID-19 patients found edema as well as lymphocyte and histiocytic infiltration, consistent with viral orchitis. [123]. These samples were also characterized by damage to the seminiferous tubules with a significant effect on Sertoli cells, as well as a reduced number of Leydig cells. In a separate study, germ cell damage was more pronounced despite similar Sertoli cell counts between SARS-CoV-2-infected individuals and uninfected controls, which represents a more direct relationship between infection and fertility. [124]. The extent to which SARS-CoV-2 may persist in the male reproductive tract is still being investigated. Although SARS-CoV-2 has been detected in human semen, it is questionable whether this is a true testicular infection or a consequence of a disrupted blood-epididymal / deferent barrier. [125,126].
Our knowledge of COVID-19 in the female reproductive system is still limited by the amount of literature and sample size of existing studies. Nevertheless, an understanding of the extent of SARS-CoV-2 in the female reproductive tract is essential to recognize any adverse effects on fertility. ACE2 is expressed in the ovaries, oocytes, and uterus, but limited coexpression of proteases such as TMPRSS2 and cathepsins L and B with ACE2 raises questions about the likelihood of ovarian / uterine infection. [127,128]. While in one study of 35 women diagnosed with COVID-19, SARS-CoV-2 was not detected in vaginal fluid or in exfoliated cervical cells, in a case study from Italy, SARS-CoV-2 was detected in vaginal fluid by RT-PCR ( Ct 37.2 on day 7 and Ct 32.9 on day 20 from the onset of symptoms), suggesting that infection of the female reproductive system is possible [129,130].
5.7. Immunological response
FIP is classically characterized as an immune-mediated disease based on early observations of complement and immunoglobulin circulation, even in the form of immune complexes. [131]. Components of type III and IV immune responses have been described [132]. Vasculitis and vasculitis-like lesions are thought to play a role in COVID-19 systemic complications that cannot be explained by direct organ infection, such as microthrombosis in the brain, kidney, spleen and liver. [133]. One type III hypersensitivity report has been identified in the COVID-19 literature [134]; however, immune complexes do not appear to play an important role in COVID-19 pathology. The mechanism of viral clearance and the inflammatory effects of the immune response are important areas of study for both FIP and COVID-19. Previous work investigating SARS-CoV has demonstrated the need for CD4 + T cells for virus clearance [135,136]. T-cell depletion was a recognized consequence of FCoV and was observed to be associated with more severe cases of COVID-19 [137,138,139]. In addition, FIP reduces both regulatory T cells and NK cells in the blood, mesenteric lymph nodes and spleen. [140]. High levels of IL-6 have previously been demonstrated in FIP ascites [50], and similarly, elevated levels of IL-6 appear to be related to disease severity and outcome in patients with COVID-19. [141]. The cytokine storm, characterized by overexpression of inflammatory cytokines, has been implicated in the pathogenesis of both infections. In FIP, this pathology is associated with monocyte and macrophage activation, while in COVID-19, the association with macrophages and monocytes is less clear. [142]. When considering the balance between cell-mediated immunity and humoral immunity, early reports suggested a link to strong humoral immunity leading to FIP [143]. However, humoral immunity may play a more beneficial role in patients with COVID-19 [144], especially given the potential clinical benefit of convalescent plasma / serum [145].
During the development of the SARS-CoV-2 vaccine, the antibody-dependent infection enhancement process (ADE), in which virus-antibody complexes enhance infection, was particularly important. FIPV has been shown to exhibit ADE in the presence of anti-FIPV antibodies [146]. This increase in infection appears to be serotype specific, with passive immunization of cats against FIPV type I or type II leading to ADE only after challenge with the same serotype for which the immunization was performed. [147]. As a result, ADE is a major challenge for the development of FIP vaccines. In diseases caused by human coronaviruses, ADE has yet to be fully understood. SARS-CoV has been found to have higher concentrations of anti-spike antibodies having a higher neutralizing effect, while more dilute concentrations are thought to contribute to ADE in vitro. [148]. In SARS-CoV-2, ADE was observed in monocyte lines but was not related to the regulation of pro-inflammatory cytokines [149]. Spike protein sequence modeling has identified possible ADE mechanisms that involve interaction with Fc receptors on monocytes and adipocytes [150]. If ADE played a role in SARS-CoV-2, the most likely mechanism would be overactivation of the immune cascade through Fc-mediated innate immune cell activation. [151,152]. There is currently insufficient evidence to suggest an ADE with the pathogenesis of SARS-CoV-2 and further research is needed to assess the true extent of the risk.
6. Molecular similarities between FCoV and SARS-CoV-2 spike proteins
Spike protein is a major factor in tissue and cell tropism and binds the cell receptor [153]. It is now well known that SARS-CoV-2 binds angiotensin converting enzyme-2 (ACE-2) as a primary receptor, a common feature of SARS-CoV. There are other binding partners for SARS-CoV-2, including heparan sulfate as a non-specific binding and neuropilin-1 (NRP-1), which may cause viral tropism for the olfactory and central nervous systems. [154,155]. In contrast, most alpha-virus viruses, including FCoV type II, use aminopeptidase (APN) virus entry. [9,153,156]. The receptor for FCoV type I has yet to be elucidated. The spike protein also mediates membrane fusion, which is activated by a complex process controlled by host cell proteases. [153]. While type I FCoV has two protease cleavage activation sites, designated S1 / S2 and S2 ′, type II FCoV has only one cleavage activation site (S2 ′). [10]. In comparison, SARS-CoV-2 is similar to FCoV-1 (and currently unique to SARS-related viruses) in that it has two identified cleavage sites (S1 / S2 and S2 ′), the first of which, the furin cleavage site or FCS is considered a significant factor in pandemic spread [157,158,159]. In both cases, the presence of S1 / S2 cleavage sites distinguishes FCoV-1 and SARS-CoV-2 from their close relatives. The importance of the cleavage activation site appears to be directly related to the proteases required for viral infection, and thus to another component of tissue tropism. In FCoV type I, the transition from FECV to macrophage-tropical FIPV was first demonstrated by amino acid substitutions at the S1 / S2 cleavage site in FIP-confirmed pathological specimens that are thought to reduce proteolytic priming of furin by similar proteases prior to S2-mediated fusion process activation. [72,160,161]. In SARS-CoV-2, TMPRSS-2 or other trypsin-like related proteases are a major activator of fusion and entry at S2 ′ [162] (Table 1), wherein the furin-like proteases prim the tip and S1 / S2 [163] and in particular have been shown to be rapidly regulated after adaptation to Vero E6 cells in culture and possibly also in extrapulmonary human tissues [164]. Thus, there appear to be remarkable similarities in host cell adaptation between the two viruses.
Virus
Group
Receptor
Consensus sequence S1 / S2 in circulating viruses
Consensus sequence S2 ′ in circulating viruses
SARS-CoV-2
Beta-coronavirus
ACE2
SPRRAR | S (* SHRRAR | S and SRRRAR | S)
SKPSKR | S
FCoV-1
Alphacoronavirus ("clade A")
unknown
SRRSRR | S (in FECV; mutated in FIPV)
KR | S
FCoV-2
Alphacoronavirus ("clade B")
APN
absent
YRKR | S
Table 1 Summary of SARS-CoV-2 and two FCoV serotypes. The coronavirus spike glycoprotein, mediated by proteolytic cleavage, is a major driver of cell receptor binding and membrane fusion. The Taxonomic classification, host receptor, and amino acid sequences of the proteolytic cleavage site S1 / S2 and S2 ′ are summarized below. *, Replaced in common variants.
7. Prevention and treatment: From social withdrawal to vaccines
Until now, the role of public health / public health measures has been a major driver in mitigating both the spread of FCoV and SARS-CoV-2. [3,31,165,166]. In this regard, many measures have been introduced for the affected population to reduce social distance, including orders to stay at home, the closure of unnecessary establishments and restrictions on public assemblies. [167]. Although not referred to as social distances, similar methods are often introduced or recommended in cat populations. [3]. Dreschler et al. summarize the methods that have been recommended in cat populations, especially in a multi-cat environment, including reducing the number of cats per room, frequent cage cleaning, and grouping cats according to excretion and / or serological status. [168]. Dreschler argues that quarantining cats exposed to FCoV / FIPV to limit the spread of FCoV in the population is neither effective nor beneficial given the likelihood of widespread FCoV infection in a multi-cat environment as well as the months required for development (and uncertainty in development) FIP. In contrast, quarantine people exposed to SARS-CoV-2 has the potential to reduce the spread of the disease and mortality [169]. Regardless of the extent of grouping or separation, the social difficulties caused by separation must be carefully considered for both cats and humans. In the case of cats, especially in connection with the premature weaning of their mothers, special attention must be paid in the weaning process to ensuring adequate socialization of the kittens. Similarly, in the case of COVID-19, the process of quarantine and / or isolation can be psychologically burdensome for individuals. A thorough cost-benefit analysis must often be carried out to compare the benefits of quarantine and isolation for public health with the negative psychological burden on the persons concerned in order to avoid unnecessary / ineffective quarantine. Where appropriate, justification as well as support to improve well-being should be provided [170].
Although the FIP vaccine is commercially available (Primucell), the benefit of FIP vaccination is still low. Primucell is an intranasal vaccine that uses an attenuated FIPV serotype 2 isolate. Although the FIP vaccine is commercially available (Primucell), the benefit of FIP vaccination is still low. Primucell is an intranasal vaccine that uses an attenuated FIPV serotype 2 isolate (FIPV-DF2), given in two doses 3 to 4 weeks apart to cats at least 16 weeks old. [171]. In a placebo-controlled experimental study in 138 cats, vaccinated cats did not show a significantly lower incidence of FIP compared to controls during the 12-month study period. After adjusting for FCoV titers, cats with lower antibody titers (100 or less) had a significantly lower incidence of FIP at the time of the first vaccination compared to cats with higher titers (400 or more). [172]. However, due to the high prevalence of FCoV, especially in multi-cat environments, attempts to alleviate FIP by vaccinating cats that are FCoV-naive at least 16 weeks of age may not be feasible due to the high potential for FCoV infection during the 16 weeks prior to vaccination. As a result, the American Association of Animal Hospitals and the American Association of General Practitioners for Cats do not recommend FIP vaccination. [173].
ADE remains a major concern of FIP vaccines. Several studies have attempted to reduce the incidence of FIP in experimentally infected cats with recombinant and other experimental vaccines, but ADEs have been reported repeatedly. In one placebo-controlled study in which purebred British Shorthairs and Specific Pathogen Free (SPF) Domestic Shorthairs were vaccinated with one of two recombinant FIPV type 2 (FIPV-DF2) vaccines, both candidate vaccines showed significantly reduced to no protection against challenge FIPV in non-SPF cats - with the majority of non-SPF animals showing ADE [174]. In a separate study, immunization of kittens with a vaccine virus recombined with the FIPV spike glycoprotein gene significantly shortened survival time after FIPV challenge compared to kittens immunized with wild-type vaccine virus. Importantly, low levels of neutralizing antibodies were observed in the group immunized against FIPV [175]. Concerns about ADE after FIPV immunization remain a challenging challenge in FIP prevention.
Unlike the FIP vaccination, the COVID-19 vaccines have played a more significant role in reducing the spread of the infection. Several types of vaccines have been produced that have demonstrated safety and efficacy in preventing symptomatic infection, severe disease, and death from COVID-19—including, but not limited to, mRNA vaccines (Pfizer/BioNTech and Moderna), viral vector vaccines (Janssen, AstraZeneca), and inactivated virus vaccines (Bharat Biotech, Sinovac) [176,177,178,179,180,181]. The first two vaccine platforms use the SARS-CoV-2 spike glycoprotein as the immunogen, while inactivated virus vaccines have the potential to elicit an immune response to viral components other than the spike glycoprotein. Despite the favorable safety profile of COVID-19 vaccines, adverse reactions occurred after vaccination, some of which were mediated by antibodies in analogy to ADE concerns with FIP vaccines. Thrombosis has been a documented problem, especially with AstraZeneca and Janssen vaccines. Although the exact mechanisms are being investigated, the inflammatory response is currently thought to lead to increased levels of platelet-activating antibodies, which bind to platelet factor 4 and lead to a hypercoagulable state. [182,183]. In contrast to the higher incidence of ADE in experimental FIP vaccines, the incidence of thrombotic events after administration of COVID-19 is low. [184].
In addition to the primary endpoints of vaccine studies, which focused on the prevention of symptomatic infection, serious illness, and death from COVID-19, many phase 3 vaccine studies did not address observations to assess the degree of prevention of asymptomatic infection. Beneficial efficacy against asymptomatic infection is important from a public health perspective, especially given that asymptomatic individuals can transmit COVID-19 and that routine monitoring testing is resource intensive and difficult to coordinate on a large scale. [22]An important contribution to this field is the real-world studies investigating vaccine efficacy, which show a reduced risk of SARS-CoV-2 infection, as well as a reduced viral load in vaccine “breakthrough” infections. [185,186,187,188]. Such evidence supports the use of SARS-CoV-2 vaccines as a protective measure not only against severe COVID-19, but also as a crucial contribution in the management of the disease.
8. Clinical care and therapeutic options
In 1963, when the first clinical cases of FIP were described (before understanding the viral etiology), it was found that antibiotic treatment was often tried, but it clearly did not bring any benefit. [189]. Since this first report and without an effective vaccine, a number of therapies have been tried in cats with FIP. Ribavirin, a nucleoside analogue, has previously provided promising results against FCoV in an in vitro study [190], however, when administered to cats as an experimental treatment, led to poorer results in some cases [191]. Similarly, at the onset of the COVID-19 pandemic, ribavirin was used in several doses and in combination with other drugs. [192] and a study protocol was designed to examine the benefit in human patients [193]. However, another direct-acting antiviral drug (DAA) (remdesivir), a nucleoside analogue that acts as a chain terminator and raises minor toxicity concerns compared to ribavirin, is rapidly gaining prominence in the treatment of hospitalized patients with COVID-19. . Despite initial enthusiasm, remdesivir has not been shown to be effective in patients with such diseases in robust clinical trials; however, several reports have demonstrated the clinical benefit of the related nucleoside analog GS-441524 in the treatment of cats with FIP, including effusive, non-fusive, and neurological forms of the disease. [194,195,196,197]. At the time of writing, studies on the effectiveness of remdesivir in the treatment of FIP are being conducted in Australia and the United Kingdom. Interestingly, remdesivir is a prodrug form of GS-441524 [195]. Two orally available DAAs have recently entered clinical trials for COVID-19 and are currently awaiting FDA approval; molnupiravir (MK-4482 / EIDD-2801), a modified form of ribavirin, and Paxlovid (a protease inhibitor PF-07321332 in combination with ritonavir, which improves the half-life of PF-07321332) targeting the major protease (Mpro). It is noteworthy that the active substance Paxlovid is related to GC-376 and has previously been shown to be effective in a FIP clinical study. [196]. It will be very interesting to follow the development, FDA approval and use of these DAAs in relation to the relevant diseases caused by SARS-CoV-2 and FCoV.
Due to the inflammatory nature of both FIP and COVID-19, treatment often focuses on controlling the immune response. Although cats with FIP are often given glucocorticoids in an effort to alleviate the inflammatory symptoms of the disease, the clinical benefit is negligible. [198]. The use of corticosteroids in patients with COVID-19 does not appear to be insignificant, with some studies showing negative profiles [199]. However, their administration may be beneficial in severe cases of COVID-19 through the observed reduction in mortality [200,201]. Cyclosporine, an immunosuppressive drug that is often used to prevent organ rejection in transplant patients and to treat some autoimmune diseases, has been studied in both FIP and SARS-CoV-2. An in vitro study with cyclosporin A (CsA) using FCoV type II virus showed a reduction in virus replication [202], treatment of a 14-year-old cat CsA after unsuccessful IFN treatment resulted in clinical improvement, reduction in viral load and survival of more than 260 days [203]. Although there are currently no controlled studies on the use of CsA in the treatment of patients with COVID-19, in addition to safety concerns, potential mechanisms of action have been suggested. [204,205,206]. In addition, the cyclosporin A analogue, alisporivir, has shown in vitro effects on virus replication [207], similar to evidence that cyclophilin A blockade inhibits the replication of other coronaviruses [208].
Many antibiotics have been prescribed for both FIP and COVID-19, but not for their antimicrobial properties, but rather for their anti-inflammatory effects. [198]. For example, doxycycline may have helped prolong survival in cats with FIP [209]. Whether doxycycline would be of benefit to patients with COVID-19 is not currently known, but has been suggested as a possible part of the treatment of the disease. [210].
Interferons have also been studied in the treatment of FIP without a clear link to clinical improvement [211]. In human patients with COVID-19, combination therapy with interferon-β-1b with lopinavir, ritonavir and ribavirin compared with lopinavir and ritonavir alone was associated with a reduced duration of virus excretion and improved clinical outcomes in mild to moderate cases. [212].
Monoclonal antibodies targeting components of the immune response have the potential to reduce inflammatory cytokine levels. A small study in cats experimentally infected with FIPV-1146 demonstrated the benefit of anti-TNF-α in managing the disease [213]. Tocilizumab, an anti-IL-6 monoclonal antibody, was administered to patients with COVID-19 [214]. Due to the different clinical results reported, further research is needed with Tocilizumab [215,216].
The transfer of knowledge between species will undoubtedly affect cats as well as humans and even other species. Although many compounds are effective when studied in vitro, their use in vivo can lead to different results, including toxicity. In addition, the fact that a compound may show promising results in one species does not mean that the same effect will be observed in other species, especially when comparing similar but different viruses and virus-induced diseases.
9. MIS-C and PASC
In April 2020, the UK's National Health Service issued an alert about an increased incidence of multisystem inflammatory syndrome in children - many of whom tested positive for COVID-19 [217]. As the pandemic progressed, studies from other countries examining this inflammatory condition provided more detail towards a clinical understanding of what is now referred to as MIS-C, a rare presentation of COVID-19 in pediatric patients. MIS-C includes several organ systems. Cardiovascular dysregulation in MIS-C is often observed in the form of ventricular dysfunction, pericardial effusion and coronary artery aneurysms [218,219]. Gastrointestinal symptoms mimic appendicitis and include abdominal pain, vomiting, and diarrhea. Terminal ileitis is a common finding on imaging tests [220]. Many patients also experience neurocognitive symptoms, including headache and confusion. More serious neurological complications, including encephalopathy and stroke, are less common [218,221].
One area of significant clinical overlap between FIP and COVID-19 is the rare inflammatory manifestation of SARS-CoV-2 infection – multisystem inflammatory syndrome in children (MIS-C). MIS-C is seen in the pediatric population, just as FIP commonly affects young cats [43]. Like FIP, MIS-C has a systemic presentation involving multiple organ systems—including gastrointestinal, cardiovascular, and hematologic abnormalities [222]. As with the wet form of FIP, pleural effusions and ascites occur in MIS-C. [223]. Both syndromes also show overlap in vascular pathology. FIP shows granulomatous vasculitis, which overlaps with Kawasaki vascular syndrome observed in MIS-C [224]. MIS-C is thought to be a post-infectious disease associated with a previous SARS-CoV-2 infection [223,225]. FIP also has a delayed onset after the first exposure to FCoV and occurs only in a small subset of cases. Although cats with FIP can still shed FCoV in their feces, the mutations associated with the biotype switch from FECV to FIPV are thought not to be transmissible—supporting some degree of similarity in the limited infectious range of both FIP and MIS-C.
A post-acute sequelae of COVID-19 (PASC) condition has recently been defined, which includes memory loss, gastrointestinal distress, fatigue, anosmia, dyspnea, etc. and is more commonly referred to as “long-term COVID.” Along with MIS-C, PASC is a very active subject of research, summarized by others [226], and together represent an excellent starting point for the use of feline medicine as a model of coronavirus-induced pathogenesis, possibly in an unexpected way [224].
10. SARS-CoV-2 infection in cats
Cats have now become widespread hosts for SARS-CoV-2 infections, in part due to the relative similarity of human and feline ACE2 receptors. Following cases reported in Hong Kong and Belgium in March 2020, the most notable early natural infection occurred at the Bronx Zoo in New York City, USA. In April, four tigers and three lions showed mild respiratory symptoms from their breeders, and SARS-CoV-2 was detected by PCR and sequencing. [227]. Consequently, infection of both domesticated and non-domesticated cats has become relatively common in cases where owners and caregivers are positive for SARS-CoV-2. From a clinical point of view, SARS-CoV-2 infection in cats is considered to be predominantly asymptomatic, with some animals showing mild respiratory symptoms. [228,229,230]. In general, severe respiratory symptoms do not appear to occur in cats, although severe respiratory distress may in some cases be related to the underlying hypertrophic cardiomyopathy (HCM) of cats. [94]. An increased incidence of myocarditis in dogs and cats has also been reported in the United Kingdom, associated with a sharp increase in variant B.1.1.7 (Alpha). [231]. There is a clear need for further studies in this area, as well as possible links between coronavirus infections in cats and multisystem inflammatory syndrome in children (MIS-C), which, as mentioned above, is a rare manifestation of COVID-19.
Laboratory animal studies have also been key to understanding SARS-CoV-2 infection in cats, which are very susceptible to infection by oronasal challenge. Experimentally infected cats showed mild respiratory symptoms or asymptomatic infection, virus shedding, virus-to-cat virus transmission, and a strong neutralizing antibody response. Recent studies have shown that long-term immunity exists after re-infection of cats, but cats may develop long-term consequences, including persistence of inflammation and other lung lesions. [232]. Overall, as with SARS-CoV in 2003, cats in particular can be an important source of information on the pathogenesis and immune responses elicited by SARS-CoV-2.
Thanks
Thanks to Annette Choi for helping with Figure 1 and all Whittaker Lab members for helpful discussions during the preparation of this manuscript.
Author shares
All authors have contributed to this article. All authors have read and agreed to the published version of the manuscript.
Financing
The work in the author's laboratory is partly funded by research grants from the National Institutes of Health, the EveryCat Foundation and the Cornell Feline Health Center. AES was supported by the NIH Comparative Medicine Training Program T32OD011000. Studies on FIP are also supported by the Michael Zemsky Fund for Cat Diseases.
Conflict of interests
The authors do not indicate any conflict of interest.
Footnotes
Publisher Note: The MDPI remains neutral in terms of jurisdictional claims in published maps and institutional affiliation.
Holzworth, J. Some Important Disorders of Cats. Cornell Vet.1963, 53, 157–160. [Google Scholar]
Hartmann, K. Feline infectious peritonitis. Vet. Clin. U.S. Small Anim. Pract.2005, 35, 39–79. [Google Scholar] [CrossRef] [PubMed]
Chen, J .; Qi, T .; Liu, L .; Ling, Y .; Qian, Z .; Li, T .; Li, F .; Xu, Q .; Zhang, Y .; Xu, S .; et al. Clinical progression of patients with COVID-19 in Shanghai, China. J. Infect.2020, 80, e1 – e6. [Google Scholar] [CrossRef] [PubMed]
Coronaviridae Study Group of the International Committee on Taxonomy of Virus. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol.2020, 5, 536–544. [Google Scholar] [CrossRef]
Perlman, SMK Coronaviruses, Including Severe Acute Respiratory Syndrome (SARS) and Middle East Respiratory Syndrome (MERS). In Mandell, Douglas, and Bennett's Principles and Practice of Infectious Disease, 9th ed .; Bennett, JE, Dolin, R., Blaser, MJ, Eds .; Elsevier: Amsterdam, The Netherlands, 2020; pp. 2072–2080. [Google Scholar]
Abdul-Rasool, S .; Fielding, BC Understanding Human Coronavirus HCoV-NL63. Open Virol. J.2010, 4, 76–84. [Google Scholar] [CrossRef]
Letko, M .; Marzi, A .; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol.2020, 5, 562–569. [Google Scholar] [CrossRef]
Rottier, PJ; Nakamura, K .; Schellen, P .; Volders, H .; Haijema, BJ Acquisition of macrophage tropism during the pathogenesis of feline infectious peritonitis is determined by mutations in the feline coronavirus spike protein. J. Virol.2005, 79, 14122–14130. [Google Scholar] [CrossRef]
Jaimes, JA; Millet, JK; Stout, AE; Andre, NM; Whittaker, GR A Tale of Two Viruses: The Distinct Spike Glycoproteins of Feline Coronaviruses. Viruses2020, 12, 83. [Google Scholar] [CrossRef]
Tresnan, DB; Levis, R .; Holmes, KV Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I. J. Virol.1996, 70, 8669–8674. [Google Scholar] [CrossRef]
Benetka, V .; Kubber-Heiss, A .; Kolodziejek, J .; Nowotny, N .; Hofmann-Parisot, M .; Mostl, K. Prevalence of feline coronavirus types I and II in cats with histopathologically verified feline infectious peritonitis. Vet. Microbiol.2004, 99, 31–42. [Google Scholar] [CrossRef]
Iwasaki, M .; Saito, J .; Zhao, H .; Sakamoto, A .; Hirota, K .; Ma, D. Inflammation Triggered by SARS-CoV-2 and ACE2 Augment Drives Multiple Organ Failure of Severe COVID-19: Molecular Mechanisms and Implications. Inflammation2021, 44, 13–34. [Google Scholar] [CrossRef]
Xiao, F .; Sun, J .; Xu, Y .; Li, F .; Huang, X .; Li, H.; Zhao, J .; Huang, J .; Zhao, J. Infectious SARS-CoV-2 in Feces of Patient with Severe COVID-19. Emerg. Infect. Dis.2020, 26, 1920–1922. [Google Scholar] [CrossRef]
Sykes, JE Feline Coronavirus Infection. In Canine and Feline Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2014; pp. 195–208. [Google Scholar] [CrossRef]
Pedersen, NC A review of feline infectious peritonitis virus infection: 1963-2008. J. Feline Med. Surg.2009, 11, 225–258. [Google Scholar] [CrossRef] [PubMed]
Pedersen, NC; Liu, H .; Scarlett, J .; Leutenegger, CM; Golovko, L .; Kennedy, H .; Kamal, FM Feline infectious peritonitis: Role of the feline coronavirus 3c gene in intestinal tropism and pathogenicity based upon isolates from resident and adopted shelter cats. Virus Res.2012, 165, 17–28. [Google Scholar] [CrossRef] [PubMed]
Brown, MA Genetic determinants of pathogenesis by feline infectious peritonitis virus. Vet. Immunol. Immunopathol.2011, 143, 265–268. [Google Scholar] [CrossRef] [PubMed]
Healey, EA; Andre, NM; Miller, AD; Whittaker, GR; Berliner, EA An outbreak of FIP in a cohort of shelter-housed cats: Molecular analysis of the feline coronavirus S1 / S2 cleavage site consistent with a “circulating virulent-avirulent” theory of FIP pathogenesis. J. Feline Med. Surg. Open Rep.2022, 8, 20551169221074226. [Google Scholar] [CrossRef]
Cheung, CCL; Goh, D .; Lim, X .; Tien, TZ; Lim, JCT; Lee, JN; Tan, B .; Tay, ZEA; Wan, WY; Chen, EX; et al. Residual SARS-CoV-2 viral antigens detected in GI and hepatic tissues from five recovered patients with COVID-19. Good2022, 71, 226–229. [Google Scholar] [CrossRef] [PubMed]
Kipar, A .; Meli, ML; Baptiste, KE; Bowker, LJ; Lutz, H. Sites of feline coronavirus persistence in healthy cats. J. Gen. Virol.2010, 91, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
Rothe, C .; Schunk, M .; Sothmann, P .; Bretzel, G .; Froeschl, G .; Wallrauch, C .; Zimmer, T .; Thiel, V .; Janke, C .; Guggemos, W .; et al. Transmission of 2019-nCoV Infection from an Asymptomatic Contact in Germany. N. Engl. J. Med.2020, 382, 970–971. [Google Scholar] [CrossRef]
Bai, Y .; Yao, L .; Wei, T .; Tian, F.; Jin, DY; Chen, L .; Wang, M. Presumed Asymptomatic Carrier Transmission of COVID-19. JAMA2020, 323, 1406–1407. [Google Scholar] [CrossRef] [PubMed]
Hu, Z .; Song, C .; Xu, C .; Jin, G .; Chen, Y .; Xu, X .; Ma, H .; Chen, W .; Lin, Y .; Zheng, Y .; et al. Clinical characteristics of 24 asymptomatic infections with COVID-19 screened among close contacts in Nanjing, China. Sci. China Life Sci.2020, 63, 706–711. [Google Scholar] [CrossRef]
Jin, YH; Cai, L .; Cheng, ZS; Cheng, H .; Deng, T .; Fan, YP; Fang, C .; Huang, D .; Huang, LQ; Huang, Q .; et al. A rapid advice guideline for the diagnosis and treatment of 2019 novel coronavirus (2019-nCoV) infected pneumonia (standard version). Mil Med. Res.2020, 7, 4. [Google Scholar] [CrossRef] [PubMed]
Wolfe, LG; Griesemer, RA Feline infectious peritonitis: Review of gross and histopathologic lesions. J. Am. Vet. Med. Assoc.1971, 158 (Suppl S2), 987. [Google Scholar]
Wege, H .; Siddell, S .; ter Meulen, V. The biology and pathogenesis of coronaviruses. Curr. Top. Microbiol. Immunol.1982, 99, 165–200. [Google Scholar] [CrossRef]
Hardy, WD, Jr .; Hurvitz, AI Feline infectious peritonitis: Experimental studies. J. Am. Vet. Med. Assoc.1971, 158 (Suppl S2), 994. [Google Scholar]
Robison, RL; Holzworth, J .; Gilmore, CE Naturally occurring feline infectious peritonitis: Signs and clinical diagnosis. J. Am. Vet. Med. Assoc.1971, 158 (Suppl. 2), 981–986. [Google Scholar]
Sherding, R. Feline Infectious Peritonitis (Feline Coronavirus). Saunders Man. Small Anim. Pract.2006, 132–143. [Google Scholar] [CrossRef]
Addie, DD; Jarrett, O. A study of naturally occurring feline coronavirus infections in kittens. Vet. Rec.1992, 130, 133–137. [Google Scholar] [CrossRef]
Lutz, H .; Gut, M .; Leutenegger, CM; Schiller, I .; Wiseman, A .; Meli, M. Kinetics of FCoV infection in kittens born in catteries of high risk for FIP under different rearing conditions. In Proceedings of the Second International Feline Coronavirus / Feline Infectious Peritonitis Symposium, Glasgow, Scotland, 4–7 August 2002. [Google Scholar]
Addie, DD; Paltrinieri, S .; Pedersen, NC Secong international feline coronavirus / feline infectious peritonitis, symposium Recommendations from workshops of the second international feline coronavirus / feline infectious peritonitis symposium. J Feline Med. Surg.2004, 6, 125–130. [Google Scholar] [CrossRef]
Vivanti, AJ; Vauloup-Fellous, C .; Prevot, S .; Zupan, V .; Suffee, C .; Do Cao, J .; Benachi, A .; De Luca, D. Transplacental transmission of SARS-CoV-2 infection. Nat. Commun.2020, 11, 3572. [Google Scholar] [CrossRef]
Shende, P .; Gaikwad, P .; Gandhewar, M .; Ukey, P .; Bhide, A .; Patel, V .; Bhagat, S .; Bhor, V .; Mahale, S .; Gajbhiye, R .; et al. Persistence of SARS-CoV-2 in the first trimester of the placenta leading to transplacental transmission and fetal demise from an asymptomatic mother. Hum. Reprod.2021, 36, 899–906. [Google Scholar] [CrossRef]
Fenizia, C .; Biasin, M .; Cetin, I .; Vergani, P .; Mileto, D .; Spinillo, A .; Gismondo, MR; Perotti, F .; Callegari, C .; Mancon, A .; et al. Analysis of SARS-CoV-2 vertical transmission during pregnancy. Nat. Commun.2020, 11, 5128. [Google Scholar] [CrossRef]
Raschetti, R .; Vivanti, AJ; Vauloup-Fellous, C .; Loi, B .; Benachi, A .; De Luca, D. Synthesis and systematic review of reported neonatal SARS-CoV-2 infections. Nat. Commun.2020, 11, 5164. [Google Scholar] [CrossRef]
Lovato, A .; de Filippis, C. Clinical Presentation of COVID-19: A Systematic Review Focusing on Upper Airway Symptoms. Ear Nose Throat J.2020, 99, 569–576. [Google Scholar] [CrossRef] [PubMed]
Wee, LE; Chan, YFZ; Teo, NWY; Cherng, BPZ; Thien, SY; Wong, HM; Wijaya, L .; Toh, ST; Tan, TT The role of self-reported olfactory and gustatory dysfunction as a screening criterion for suspected COVID-19. Eur. Arch. Otorhinolaryngol.2020, 277, 2389–2390. [Google Scholar] [CrossRef]
Peckham, H .; de Gruijter, NM; Raine, C .; Radziszewska, A .; Ciurtin, C .; Wedderburn, LR; Rosser, EC; Webb, K .; Deakin, CT Male sex identified by global COVID-19 meta-analysis as a risk factor for death and ITU admission. Nat. Commun.2020, 11, 6317. [Google Scholar] [CrossRef] [PubMed]
Vahidy, FS; Pan, AP; Ahnstedt, H .; Munshi, Y .; Choi, HA; Tiruneh, Y .; Nasir, K .; Kash, BA; Andrieni, JD; McCullough, LD Sex differences in susceptibility, severity, and outcomes of coronavirus disease 2019: Cross-sectional analysis from a diverse US metropolitan area. PLoS ONE2021, 16, e0245556. [Google Scholar] [CrossRef] [PubMed]
Riemer, F.; Kuehner, KA; Ritz, S.; Sauter-Louis, C.; Hartmann, K. Clinical and laboratory features of cats with feline infectious peritonitis–a retrospective study of 231 confirmed cases (2000–2010). J. Feline Med. Surg.2016, 18, 348–356. [Google Scholar] [CrossRef]
Hambali, NL; Mohd Noh, M .; Paramasivam, S .; Chua, TH; Hayati, F .; Payus, AO; Tee, TY; Rosli, KT; Abd Rachman Isnadi, MF; Manin, BO A Non-severe Coronavirus Disease 2019 Patient With Persistently High Interleukin-6 Level. Front. Public Health2020, 8, 584552. [Google Scholar] [CrossRef]
August, JR Feline infectious peritonitis. An immune-mediated coronaviral vasculitis. Vet. Clin. U.S. Small Anim. Pract.1984, 14, 971–984. [Google Scholar] [CrossRef]
Hayashi, T .; Goto, N .; Takahashi, R .; Fujiwara, K. Systemic vascular lesions in feline infectious peritonitis. Nihon Juigaku Zasshi1977, 39, 365–377. [Google Scholar] [CrossRef]
Stout, AE; Andre, NM; Zimmerberg, J .; Baker, SC; Whittaker, GR Coronaviruses as a cause of vascular disease: A comparative medicine approach. eCommons2021. [Google Scholar] [CrossRef]
Varga, Z .; Flammer, AJ; Steiger, P .; Haberecker, M .; Andermatt, R .; Zinkernagel, AS; Mehra, MR; Schuepbach, RA; Ruschitzka, F .; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet2020, 395, 1417–1418. [Google Scholar] [CrossRef]
Becker, RC COVID-19-associated vasculitis and vasculopathy. J. Thromb. Thrombolysis2020, 50, 499–511. [Google Scholar] [CrossRef]
Goitsuka, R .; Ohashi, T .; Ono, K .; Yasukawa, K .; Koishibara, Y .; Fukui, H .; Ohsugi, Y .; Hasegawa, A. IL-6 activity in feline infectious peritonitis. J. Immunol.1990, 144, 2599–2603. [Google Scholar] [PubMed]
Malbon, AJ; Fonfara, S .; Meli, ML; Hahn, S .; Egberink, H .; Kipar, A. Feline Infectious Peritonitis as a Systemic Inflammatory Disease: Contribution of Liver and Heart to the Pathogenesis. Viruses2019, 11, 1144. [Google Scholar] [CrossRef]
Mestrinho, LA; Rosa, R .; Ramalho, P .; Branco, V .; Iglesias, L .; Pissarra, H .; Duarte, A .; Niza, M. A pilot study to evaluate the serum Alpha-1 acid glycoprotein response in cats suffering from feline chronic gingivostomatitis. BMC Vet. Res.2020, 16, 390. [Google Scholar] [CrossRef] [PubMed]
Selting, KA; Ogilvie, GK; Lana, SE; Fettman, MJ; Mitchener, KL; Hansen, RA; Richardson, KL; Walton, JA; Scherk, MA Serum alhpa 1-acid glycoprotein concentrations in healthy and tumor-bearing cats. J. Vet. Intern. Med.2000, 14, 503–506. [Google Scholar] [CrossRef]
Giordano, A .; Spagnolo, V .; Colombo, A .; Paltrinieri, S. Changes in some acute phase protein and immunoglobulin concentrations in cats affected by feline infectious peritonitis or exposed to feline coronavirus infection. Vet. J.2004, 167, 38–44. [Google Scholar] [CrossRef]
Hazuchova, K .; Held, S .; Neiger, R. Usefulness of acute phase proteins in differentiating between feline infectious peritonitis and other diseases in cats with body cavity effusions. J. Feline Med. Surg.2017, 19, 809–816. [Google Scholar] [CrossRef] [PubMed]
Li, H.; Xiang, X .; Ren, H.; Xu, L .; Zhao, L .; Chen, X .; Long, H.; Wang, Q .; Wu, Q. Serum Amyloid A is a biomarker of severe Coronavirus Disease and poor prognosis. J. Infect.2020, 80, 646–655. [Google Scholar] [CrossRef] [PubMed]
Zinellu, A .; Paliogiannis, P .; Carru, C .; Mangoni, AA Serum amyloid A concentrations, COVID-19 severity and mortality: An updated systematic review and meta-analysis. Int. J. Infect. Dis.2021, 105, 668–674. [Google Scholar] [CrossRef] [PubMed]
Nehring, SM; Goyal, A .; Bansal, P .; Patel, BC C Reactive Protein; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
Vanderschueren, S .; Deeren, D .; Knockaert, DC; Bobbaers, H .; Bossuyt, X .; Peetermans, W. Extremely elevated C-reactive protein. Eur. J. Intern. Med.2006, 17, 430–433. [Google Scholar] [CrossRef]
Yang, M .; Chen, X .; Xu, Y. A Retrospective Study of the C-Reactive Protein to Lymphocyte Ratio and Disease Severity in 108 Patients with Early COVID-19 Pneumonia from January to March 2020 in Wuhan, China. Med. Sci. Monit.2020, 26, e926393. [Google Scholar] [CrossRef]
Liu, F .; Li, L .; Xu, M .; Wu, J .; Luo, D .; Zhu, Y .; Li, B .; Song, X .; Zhou, X. Prognostic value of interleukin-6, C-reactive protein, and procalcitonin in patients with COVID-19. J. Clin. Virol.2020, 127, 104370. [Google Scholar] [CrossRef]
Sharifpour, M .; Rangaraju, S .; Liu, M .; Alabyad, D .; Nahab, FB; Creel-Bulos, CM; Jabaley, CS; Emory, C.-Q .; Clinical Research, C. C-Reactive protein as a prognostic indicator in hospitalized patients with COVID-19. PLoS ONE2020, 15, e0242400. [Google Scholar] [CrossRef]
Leisman, DE; Ronner, L .; Pinotti, R .; Taylor, MD; Sinha, P .; Calfee, CS; Hirayama, AV; Mastroiani, F.; Turtle, CJ; Harhay, MO; et al. Cytokine elevation in severe and critical COVID-19: A rapid systematic review, meta-analysis, and comparison with other inflammatory syndromes. Lancet Respir. Med.2020, 8, 1233–1244. [Google Scholar] [CrossRef]
Adam, SS; Key, NS; Greenberg, CS D-dimer antigen: Current concepts and future prospects. Blood2009, 113, 2878–2887. [Google Scholar] [CrossRef]
Wichmann, D .; Sperhake, JP; Lutgehetmann, M .; Steurer, S .; Edler, C .; Heinemann, A .; Heinrich, F .; Mushumba, H .; Kniep, I .; Schroder, AS; et al. Autopsy Findings and Venous Thromboembolism in Patients With COVID-19: A Prospective Cohort Study. Ann. Intern. Med.2020, 173, 268–277. [Google Scholar] [CrossRef]
Ackermann, M .; Verleden, SE; Kuehnel, M .; Haverich, A .; Welte, T .; Laenger, F .; Vanstapel, A .; Werlein, C .; Stark, H .; Tzankov, A .; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. N. Engl. J. Med.2020, 383, 120–128. [Google Scholar] [CrossRef] [PubMed]
Yu, HH; Qin, C .; Chen, M .; Wang, W .; Tian, DS D-dimer level is associated with the severity of COVID-19. Thromb. Res.2020, 195, 219–225. [Google Scholar] [CrossRef] [PubMed]
Kermali, M .; Khalsa, RK; Pillai, K .; Ismail, Z .; Harky, A. The role of biomarkers in diagnosis of COVID-19-A systematic review. Life Sci.2020, 254, 117788. [Google Scholar] [CrossRef] [PubMed]
Tholen, I .; Weingart, C .; Kohn, B. Concentration of D-dimers in healthy cats and sick cats with and without disseminated intravascular coagulation (DIC). J. Feline Med. Surg.2009, 11, 842–846. [Google Scholar] [CrossRef] [PubMed]
Marioni-Henry, K .; Vite, CH; Newton, AL; Van Winkle, TJ Prevalence of diseases of the spinal cord of cats. J. Vet. Intern. Med.2004, 18, 851–858. [Google Scholar] [CrossRef] [PubMed]
Andre, NM; Cossic, B .; Davies, E .; Miller, AD; Whittaker, GR Distinct mutation in the feline coronavirus spike protein cleavage activation site in a cat with feline infectious peritonitis-associated meningoencephalomyelitis. JFMS Open Rep.2019, 5, 2055116919856103. [Google Scholar] [CrossRef]
Diaz, JV; Poma, R. Diagnosis and clinical signs of feline infectious peritonitis in the central nervous system. Can. Vet. J.2009, 50, 1091–1093. [Google Scholar]
Crawford, AH; Stoll, AL; Sanchez-Masian, D .; Shea, A .; Michaels, J .; Fraser, AR; Beltran, E. Clinicopathologic Features and Magnetic Resonance Imaging Findings in 24 Cats With Histopathologically Confirmed Neurologic Feline Infectious Peritonitis. J. Vet. Intern. Med.2017, 31, 1477–1486. [Google Scholar] [CrossRef]
Zhou, L .; Zhang, M .; Wang, J .; Gao, J. Sars-Cov-2: Underestimated damage to nervous system. Travel Med. Infect. Dis.2020, 101642. [Google Scholar] [CrossRef]
Asadi-Pooya, AA; Simani, L. Central nervous system manifestations of COVID-19: A systematic review. J. Neurol. Sci.2020, 413, 116832. [Google Scholar] [CrossRef] [PubMed]
Matschke, J .; Lutgehetmann, M .; Hagel, C .; Sperhake, JP; Schroder, AS; Edler, C .; Mushumba, H .; Fitzek, A .; Allweiss, L .; Dandri, M .; et al. Neuropathology of patients with COVID-19 in Germany: A post-mortem case series. Lancet Neurol.2020, 19, 919–929. [Google Scholar] [CrossRef]
Paniz-Mondolfi, A .; Bryce, C .; Grimes, Z .; Gordon, RE; Reidy, J .; Lednicky, J .; Sordillo, EM; Fowkes, M. Central nervous system involvement by severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). J. Med. Virol.2020, 92, 699–702. [Google Scholar] [CrossRef] [PubMed]
Lee, MH; Perl, DP; Nair, G .; Li, W .; Maric, D .; Murray, H .; Dodd, SJ; Koretsky, AP; Watts, JA; Cheung, V .; et al. Microvascular Injury in the Brains of Patients with Covid-19. N. Engl. J. Med.2021, 384, 481–483. [Google Scholar] [CrossRef] [PubMed]
Andrew, SE Feline infectious peritonitis. Vet. Clin. U.S. Small Anim. Pract.2000, 30, 987–1000. [Google Scholar] [CrossRef]
Cannon, MJ; Silkstone, MA; Kipar, AM Cutaneous lesions associated with coronavirus-induced vasculitis in a cat with feline infectious peritonitis and concurrent feline immunodeficiency virus infection. J. Feline Med. Surg.2005, 7, 233–236. [Google Scholar] [CrossRef] [PubMed]
Hok, K. Demonstration of feline corona virus (FCV) antigen in organs of cats suspected of feline infectious peritonitis (FIP) disease. APMIS1990, 98, 659–664. [Google Scholar] [CrossRef] [PubMed]
Wu, P .; Duan, F .; Luo, C .; Liu, Q .; Qu, X .; Liang, L .; Wu, K. Characteristics of Ocular Findings of Patients With Coronavirus Disease 2019 (COVID-19) in Hubei Province, China. JAMA Ophthalmol.2020, 138, 575–578. [Google Scholar] [CrossRef]
Mazzotta, C .; Giancipoli, E. Anterior Acute Uveitis Report in a SARS-CoV-2 Patient Managed with Adjunctive Topical Antiseptic Prophylaxis Preventing 2019-nCoV Spread Through the Ocular Surface Route. Int. Med. Case Rep. J.2020, 13, 513–520. [Google Scholar] [CrossRef] [PubMed]
Francois, J .; Collery, AS; Hayek, G .; Sot, M .; Zaidi, M .; Lhuillier, L .; Perone, JM Coronavirus Disease 2019-Associated Ocular Neuropathy With Panuveitis: A Case Report. JAMA Ophthalmol.2021, 139, 247–249. [Google Scholar] [CrossRef]
Loon, SC; Teoh, SC; Oon, LL; Se-Thoe, SY; Ling, AE; Leo, YS; Leong, HN The severe acute respiratory syndrome coronavirus in tears. Br. J. Ophthalmol.2004, 88, 861–863. [Google Scholar] [CrossRef]
Arora, R .; Goel, R .; Kumar, S .; Chhabra, M .; Saxena, S .; Manchanda, V .; Pumma, P. Evaluation of SARS-CoV-2 in Tears of Patients with Moderate to Severe COVID-19. Ophthalmology2021, 128, 494–503. [Google Scholar] [CrossRef]
Colavita, F .; Lapa, D .; Carletti, F .; Lalle, E .; Bordi, L .; Marsella, P .; Nicastri, E .; Bevilacqua, N .; Giancola, ML; Corpolongo, A .; et al. SARS-CoV-2 Isolation From Ocular Secretions of a Patient With COVID-19 in Italy With Prolonged Viral RNA Detection. Ann. Intern. Med.2020, 173, 242–243. [Google Scholar] [CrossRef] [PubMed]
Fischer, Y .; Wess, G .; Hartmann, K. Pericardial effusion in a cat with feline infectious peritonitis. Switzerland Arch. Tierheilkd.2012, 154, 27–31. [Google Scholar] [CrossRef] [PubMed]
Rush, JE; Keene, BW; Fox, PR Pericardial disease in the cat: A retrospective evaluation of 66 cases. J. Am. Anim. Hosp. Assoc.1990, 26, 39–46. [Google Scholar]
Hall, DJ; Shofer, F .; Meier, CK; Sleeper, MM Pericardial effusion in cats: A retrospective study of clinical findings and outcome in 146 cats. J. Vet. Intern. Med.2007, 21, 1002–1007. [Google Scholar] [CrossRef]
Baek, S .; Jo, J .; Song, K .; Seo, K. Recurrent Pericardial Effusion with Feline Infectious Peritonitis in a Cat. J. Vet. Clin.2017, 34, 437–440. [Google Scholar] [CrossRef]
Ernandes, MA; Cantoni, AM; Armando, F .; Corradi, A .; Ressel, L .; Tamborini, A. Feline coronavirus-associated myocarditis in a domestic longhair cat. JFMS Open Rep.2019, 5, 2055116919879256. [Google Scholar] [CrossRef]
Carvallo, FR; Martins, M .; Joshi, LR; Caserta, LC; Mitchell, PK; Cecere, T .; Hancock, S .; Goodrich, EL; Murphy, J .; Part, DG Severe SARS-CoV-2 Infection in a Cat with Hypertrophic Cardiomyopathy. Viruses2021, 13, 1510. [Google Scholar] [CrossRef]
Guo, T .; Fan, Y .; Chen, M .; Wu, X .; Zhang, L .; He, T .; Wang, H .; Wan, J .; Wang, X .; Lu, Z. Cardiovascular Implications of Fatal Outcomes of Patients With Coronavirus Disease 2019 (COVID-19). JAMA Cardiol.2020, 5, 811–818. [Google Scholar] [CrossRef]
Ruan, Q .; Yang, K .; Wang, W .; Jiang, L .; Song, J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med.2020, 46, 846–848. [Google Scholar] [CrossRef]
Inciardi, RM; Lupi, L .; Zaccone, G .; Italy, L .; Raffo, M .; Tomasoni, D .; Cani, DS; Cerini, M .; Farina, D .; Gavazzi, E .; et al. Cardiac Involvement in a Patient With Coronavirus Disease 2019 (COVID-19). JAMA Cardiol.2020, 5, 819–824. [Google Scholar] [CrossRef]
Farina, A .; Uccello, G .; Spreafico, M .; Bassanelli, G .; Savonitto, S. SARS-CoV-2 detection in the pericardial fluid of a patient with cardiac tamponade. Eur. J. Intern. Med.2020, 76, 100. [Google Scholar] [CrossRef] [PubMed]
Lindner, D .; Fitzek, A .; Brauninger, H .; Aleshcheva, G .; Edler, C .; Meissner, K .; Scherschel, K .; Kirchhof, P .; Escher, F .; Schultheiss, HP; et al. Association of Cardiac Infection With SARS-CoV-2 in Confirmed COVID-19 Autopsy Cases. JAMA Cardiol.2020, 5, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
Gunn-Moore, DA; Gruffydd-Jones, TJ; Harbor, DA Detection of feline coronaviruses by culture and reverse transcriptase-polymerase chain reaction of blood samples from healthy cats and cats with clinical feline infectious peritonitis. Vet. Microbiol.1998, 62, 193–205. [Google Scholar] [CrossRef]
Addie, DD; Jarrett, O. Use of a reverse-transcriptase polymerase chain reaction for monitoring the shedding of feline coronavirus by healthy cats. Vet. Rec.2001, 148, 649–653. [Google Scholar] [CrossRef] [PubMed]
Stranieri, A .; Scavone, D .; Paltrinieri, S .; Giordano, A .; Bonsembiante, F .; Ferro, S .; Gelain, ME; Meazzi, S .; Lauzi, S. Concordance between Histology, Immunohistochemistry, and RT-PCR in the Diagnosis of Feline Infectious Peritonitis. Pathogens2020, 9, 852. [Google Scholar] [CrossRef]
Harvey, CJ; Lopez, JW; Hendrick, MJ An uncommon intestinal manifestation of feline infectious peritonitis: 26 cases (1986–1993). J. Am. Vet. Med. Assoc.1996, 209, 1117–1120. [Google Scholar]
Xiao, F .; Tang, M .; Zheng, X .; Liu, Y .; Li, X .; Shan, H. Evidence for Gastrointestinal Infection of SARS-CoV-2. Gastroenterology2020, 158, 1831–1833. [Google Scholar] [CrossRef]
Pan, L .; Mu, M .; Yang, P .; Sun, Y .; Wang, R .; Yan, J .; Li, P .; Hu, B .; Wang, J .; Hu, C .; et al. Clinical Characteristics of COVID-19 Patients With Digestive Symptoms in Hubei, China: A Descriptive, Cross-Sectional, Multicenter Study. Am. J. Gastroenterol.2020, 115, 766–773. [Google Scholar] [CrossRef] [PubMed]
Parasa, S .; Desai, M .; Thoguluva Chandrasekar, V .; Patel, HK; Kennedy, KF; Roesch, T .; Spadaccini, M .; Colombo, M .; Gabbiadini, R .; Artifon, ELA; et al. Prevalence of Gastrointestinal Symptoms and Fecal Viral Shedding in Patients With Coronavirus Disease 2019: A Systematic Review and Meta-analysis. JAMA Netw. Open2020, 3, e2011335. [Google Scholar] [CrossRef] [PubMed]
Rokkas, T. Gastrointestinal involvement in COVID-19: A systematic review and meta-analysis. Ann. Gastroenterol.2020, 33, 355–365. [Google Scholar] [CrossRef] [PubMed]
Akin, H.; Kurt, R .; Tufan, F .; Swi, A .; Ozaras, R .; Tahan, V .; Hammoud, G. Newly Reported Studies on the Increase in Gastrointestinal Symptom Prevalence withCOVID-19 Infection: A Comprehensive Systematic Review and Meta-Analysis. Diseases2020, 8, 41. [Google Scholar] [CrossRef] [PubMed]
Chen, L .; Lou, J .; Bai, Y .; Wang, M. COVID-19 Disease With Positive Fecal and Negative Pharyngeal and Sputum Viral Tests. Am. J. Gastroenterol.2020, 115, 790. [Google Scholar] [CrossRef] [PubMed]
Wang, W .; Xu, Y .; Gao, R .; Lu, R .; Han, K .; Wu, G .; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA2020, 323, 1843–1844. [Google Scholar] [CrossRef]
Arostegui, D .; Castro, K .; Schwarz, S .; Vaidy, K .; Rabinowitz, S .; Wallach, T. Persistent SARS-CoV-2 Nucleocapsid Protein Presence in the Intestinal Epithelium of a Pediatric Patient 3 Months After Acute Infection. J. Pediatr. Gastroenterol. Nutr.2022, 3, e152. [Google Scholar] [CrossRef]
Declercq, J .; De Bosschere, H .; Schwarzkopf, I .; Declercq, L. Papular cutaneous lesions in a cat associated with feline infectious peritonitis. Vet. Dermatol.2008, 19, 255–258. [Google Scholar] [CrossRef]
Bauer, BS; Kerr, ME; Sandmeyer, LS; Grahn, BH Positive immunostaining for feline infectious peritonitis (FIP) in a Sphinx cat with cutaneous lesions and bilateral panuveitis. Vet. Ophthalmol.2013, 16 (Suppl. 1), 160–163. [Google Scholar] [CrossRef]
Redford, T .; Al-Dissi, AN Feline infectious peritonitis in a cat presented because of papular skin lesions. Can. Vet. J.2019, 60, 183–185. [Google Scholar]
Trotman, TK; Mauldin, E .; Hoffmann, V .; Del Piero, F .; Hess, RS Skin fragility syndrome in a cat with feline infectious peritonitis and hepatic lipidosis. Vet. Dermatol.2007, 18, 365–369. [Google Scholar] [CrossRef] [PubMed]
Recalcati, S. Cutaneous manifestations in COVID-19: A first perspective. J. Eur. Acad. Dermatol. Venereol.2020, 34. [Google Scholar] [CrossRef] [PubMed]
Galvan Casas, C .; Catala, A .; Carretero Hernandez, G .; Rodriguez-Jimenez, P .; Fernandez-Nieto, D .; Rodriguez-Villa Lario, A .; Navarro Fernandez, I .; Ruiz-Villaverde, R .; Falkenhain-Lopez, D .; Llamas Velasco, M .; et al. Classification of the cutaneous manifestations of COVID-19: A rapid prospective nationwide consensus study in Spain with 375 cases. Br. J. Dermatol.2020, 183, 71–77. [Google Scholar] [CrossRef] [PubMed]
Tomsitz, D .; Biedermann, T .; Brockow, K. Skin manifestations reported in association with COVID-19 infection. J. Dtsch. Dermatol. Ges.2021, 19, 530–534. [Google Scholar] [CrossRef]
Welsh, EC; Alfaro Sanchez, AB; Ortega Gutierrez, GL; Cardenas-de la Garza, JA; Cuellar-Barboza, A .; Valdes-Espinosa, RA; Pasos Estrada, AA; Miranda Aguirre, AI; Ramos-Jimenez, J .; Moreno Gonzalez, J .; et al. COVID-19 dermatological manifestations: Results from the Mexican Academy of Dermatology COVID-19 registry. Int. J. Dermatol.2021, 60, 879. [Google Scholar] [CrossRef]
Foster, RA; Caswell, JL; Rinkardt, N. Chronic fibrinous and necrotic orchitis in a cat. Can. Vet. J.1996, 37, 681–682. [Google Scholar]
Stranieri, A .; Probo, M .; Pisu, MC; Fioletti, A .; Meazzi, S .; Gelain, ME; Bonsembiante, F .; Lauzi, S .; Paltrinieri, S. Preliminary investigation on feline coronavirus presence in the reproductive tract of the tom cat as a potential route of viral transmission. J. Feline Med. Surg.2020, 22, 178–185. [Google Scholar] [CrossRef]
Evermann, JF; Baumgartener, L .; Ott, RL; Davis, EV; McKeirnan, AJ Characterization of a feline infectious peritonitis virus isolate. Vet. Pathol.1981, 18, 256–265. [Google Scholar] [CrossRef]
Yang, M .; Chen, S .; Huang, B .; Zhong, JM; Su, H.; Chen, YJ; Cao, Q .; He had.; He, J .; Li, XF; et al. Pathological Findings in the Testes of COVID-19 Patients: Clinical Implications. Eur. Urol. Focus2020, 6, 1124–1129. [Google Scholar] [CrossRef]
Ma, X .; Guan, C .; Chen, R .; Wang, Y .; Feng, S .; Wang, R .; Qu, G .; Zhao, S .; Wang, F .; Wang, X .; et al. Pathological and molecular examinations of postmortem testis biopsies reveal SARS-CoV-2 infection in the testis and spermatogenesis damage in COVID-19 patients. Cell Mol. Immunol.2021, 18, 487–489. [Google Scholar] [CrossRef]
Li, D .; Jin, M .; Bao, P .; Zhao, W .; Zhang, S. Clinical Characteristics and Results of Semen Tests Among Men With Coronavirus Disease 2019. JAMA Netw. Open2020, 3, e208292. [Google Scholar] [CrossRef] [PubMed]
Sharun, K .; Tiwari, R .; Dhama, K. SARS-CoV-2 in semen: Potential for sexual transmission in COVID-19. Int. J. Surg.2020, 84, 156–158. [Google Scholar] [CrossRef] [PubMed]
Jing, Y .; Run-Qian, L .; Hao-Ran, W .; Hao-Ran, C .; Ya-Bin, L .; Yang, G .; Fei, C. Potential influence of COVID-19 / ACE2 on the female reproductive system. Mol. Hum. Reprod.2020, 26, 367–373. [Google Scholar] [CrossRef]
Goad, J .; Rudolph, J .; Rajkovic, A. Female reproductive tract has low concentration of SARS-CoV2 receptors. PLoS ONE2020, 15, e0243959. [Google Scholar] [CrossRef] [PubMed]
Cui, P .; Chen, Z .; Wang, T .; Dai, J .; Zhang, J .; Ding, T .; Jiang, J .; Liu, J .; Zhang, C .; Shan, W .; et al. Severe acute respiratory syndrome coronavirus 2 detection in the female lower genital tract. Am. J. Obstet. Gynecol.2020, 223, 131–134. [Google Scholar] [CrossRef]
Scorzolini, L .; Corpolongo, A .; Castilletti, C .; Lalle, E .; Mariano, A .; Nicastri, E. Comment on the Potential Risks of Sexual and Vertical Transmission of COVID-19. Clin. Infect. Dis.2020, 71, 2298. [Google Scholar] [CrossRef] [PubMed]
Petersen, NC; Boyle, JF Immunologic phenomena in the effusive form of feline infectious peritonitis. Am. J. Vet. Res.1980, 41, 868–876. [Google Scholar]
Kipar, A .; May, H .; Menger, S .; Weber, M .; Leukert, W .; Reinacher, M. Morphologic features and development of granulomatous vasculitis in feline infectious peritonitis. Vet. Pathol.2005, 42, 321–330. [Google Scholar] [CrossRef]
McGonagle, D .; Bridgewood, C .; Ramanan, AV; Meaney, JFM; Watad, A. COVID-19 vasculitis and novel vasculitis mimics. Rheumatol Lancet.2021, 3, e224 – e233. [Google Scholar] [CrossRef]
Roncati, L .; Ligabue, G .; Fabbiani, L .; Malagoli, C .; Gallo, G .; Lusenti, B .; Nasillo, V .; Manenti, A .; Maiorana, A. Type 3 hypersensitivity in COVID-19 vasculitis. Clin. Immunol.2020, 217, 108487. [Google Scholar] [CrossRef]
Chen, J .; Lau, YF; Lamirande, EW; Paddock, CD; Bartlett, JH; Zaki, SR; Subbarao, K. Cellular immune responses to severe acute respiratory syndrome coronavirus (SARS-CoV) infection in senescent BALB / c mice: CD4 + T cells are important in controlling of SARS-CoV infection. J. Virol.2010, 84, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
Yasui, F .; Kohara, M .; Kitabatake, M .; Nishiwaki, T .; Fujii, H .; Tateno, C .; Yoneda, M .; Morita, K .; Matsushima, K .; Koyasu, S .; et al. Phagocytic cells contribute to the antibody-mediated elimination of pulmonary-infected SARS coronavirus. Virology2014, 454–455, 157–168. [Google Scholar] [CrossRef] [PubMed]
Chen, G .; Wu, D .; Guo, W .; Cao, Y .; Huang, D .; Wang, H .; Wang, T .; Zhang, X .; Chen, H .; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig.2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
de Groot-Mijnes, JD; van Dun, JM; van der Most, RG; de Groot, RJ Natural history of a recurrent feline coronavirus infection and the role of cellular immunity in survival and disease. J. Virol.2005, 79, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
Haagmans, BL; Egberink, HF; Horzinek, MC Apoptosis and T-cell depletion during feline infectious peritonitis. J. Virol.1996, 70, 8977–8983. [Google Scholar] [CrossRef]
Vermeulen, BL; Devriendt, B .; Olyslaegers, DA; Dedeurwaerder, A .; Desmarets, LM; Favoreel, HW; Dewerchin, HL; Nauwynck, HJ Suppression of NK cells and regulatory T lymphocytes in cats naturally infected with feline infectious peritonitis virus. Vet. Microbiol.2013, 164, 46–59. [Google Scholar] [CrossRef]
Aziz, M .; Fatima, R .; Assaly, R. Elevated interleukin-6 and severe COVID-19: A meta-analysis. J. Med. Virol.2020, 92, 2283–2285. [Google Scholar] [CrossRef]
Merad, M .; Martin, JC Author Correction: Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol.2020, 20, 448. [Google Scholar] [CrossRef]
Kai, K .; Yukimune, M .; Murata, T .; Uzuka, Y .; Canoe, M .; Matsumoto, H. Humoral immune responses of cats to feline infectious peritonitis virus infection. J. Vet. Med. Sci.1992, 54, 501–507. [Google Scholar] [CrossRef]
Ni, L .; Ye, F .; Cheng, ML; Feng, Y .; Deng, YQ; Zhao, H .; Wei, P .; Ge, J .; Gou, M .; Li, X .; et al. Detection of SARS-CoV-2-Specific Humoral and Cellular Immunity in COVID-19 Convalescent Individuals. Immunity2020, 52, 971–977. [Google Scholar] [CrossRef]
Kong, Y .; Cai, C .; Ling, L .; Zeng, L .; Wu, M .; Wu, Y .; Zhang, W .; Liu, Z. Successful treatment of a centenarian with coronavirus disease 2019 (COVID-19) using convalescent plasma. Transfus. Apher. Sci.2020, 59, 102820. [Google Scholar] [CrossRef]
Olsen, CW; Corapi, WV; Ngichabe, CK; Baines, JD; Scott, FW Monoclonal antibodies to the spike protein of feline infectious peritonitis virus mediate antibody-dependent enhancement of infection of feline macrophages. J. Virol.1992, 66, 956–965. [Google Scholar] [CrossRef]
Takano, T .; Kawakami, C .; Yamada, S .; Satoh, R .; Hohdatsu, T. Antibody-dependent enhancement occurs upon re-infection with the identical serotype virus in feline infectious peritonitis virus infection. J. Vet. Med. Sci.2008, 70, 1315–1321. [Google Scholar] [CrossRef]
Maemura, T .; Kuroda, M .; Armbrust, T .; Yamayoshi, S .; Halfmann, PJ; Kawaoka, Y. Antibody-Dependent Enhancement of SARS-CoV-2 Infection Is Mediated by the IgG Receptors FcgammaRIIA and FcgammaRIIIA but Does Not Contribute to Aberrant Cytokine Production by Macrophages. mBio2021, 12, e0198721. [Google Scholar] [CrossRef]
Ricke, DO Two Different Antibody-Dependent Enhancement (ADE) Risks for SARS-CoV-2 Antibodies. Front. Immunol.2021, 12, 640093. [Google Scholar] [CrossRef] [PubMed]
Hui, KPY; Cheung, MC; Perera, R .; Ng, KC; Bui, CHT; Ho, JCW; Ng, MMT; Kuok, DIT; Shih, KC; Tsao, SW; et al. Tropism, replication competence, and innate immune responses of the coronavirus SARS-CoV-2 in human respiratory tract and conjunctiva: An analysis in ex-vivo and in-vitro cultures. Lancet Respir. Med.2020, 8, 687–695. [Google Scholar] [CrossRef]
Li, F. Structure, Function, and Evolution of Coronavirus Spike Proteins. Annu. Rev. Virol.2016, 3, 237–261. [Google Scholar] [CrossRef] [PubMed]
Clausen, TM; Sandoval, DR; Spliid, CB; Pihl, J .; Perrett, HR; Painter, CD; Narayanan, A .; Majowicz, SA; Kwong, EM; McVicar, RN; et al. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2. Cell2020, 183, 1043–1057. [Google Scholar] [CrossRef] [PubMed]
Cantuti-Castelvetri, L .; Ojha, R .; Pedro, LD; Djannatian, M .; Franz, J .; Kuivanen, S .; van der Meer, F .; Kallio, K .; Kaya, T .; Anastasina, M .; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science2020, 370, 856–860. [Google Scholar] [CrossRef] [PubMed]
Lan, J .; Ge, J .; Yu, J .; Shan, S .; Zhou, H .; Fan, S .; Zhang, Q .; Shi, X .; Wang, Q .; Zhang, L .; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed]
Hoffmann, M .; Kleine-Weber, H .; Pohlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell2020, 78, 779–784. [Google Scholar] [CrossRef] [PubMed]
Whittaker, GR SARS-CoV-2 spike and its adaptable furin cleavage site. Lancet Microbe2021, 2, e488 – e489. [Google Scholar] [CrossRef]
Wrobel, AG; Benton, DJ; Xu, P .; Roustan, C .; Martin, SR; Rosenthal, PB; Skehel, JJ; Gamblin, SJ SARS-CoV-2 and bat RaTG13 spike glycoprotein structures inform on virus evolution and furin-cleavage effects. Nat. Struct. Mol. Biol.2020, 27, 763–767. [Google Scholar] [CrossRef] [PubMed]
Licitra, BN; Millet, JK; Regan, AD; Hamilton, BS; Rinaldi, VD; Duhamel, GE; Whittaker, GR Mutation in spike protein cleavage site and pathogenesis of feline coronavirus. Emerg. Infect. Dis.2013, 19, 1066–1073. [Google Scholar] [CrossRef]
Andre, NM; Miller, AD; Whittaker, GR Feline infectious peritonitis virus-associated rhinitis in a cat. JFMS Open Rep.2020, 6, 2055116920930582. [Google Scholar] [CrossRef]
Jaimes, JA; Millet, JK; Whittaker, GR Proteolytic Cleavage of the SARS-CoV-2 Spike Protein and the Role of the Novel S1 / S2 Site. iScience2020, 23, 101212. [Google Scholar] [CrossRef]
Tang, T .; Jaimes, JA; Bidon, MK; Straus, MR; Daniel, S .; Whittaker, GR Proteolytic Activation of SARS-CoV-2 Spike at the S1 / S2 Boundary: Potential Role of Proteases beyond Furin. ACS Infect. Dis.2021, 7, 264–272. [Google Scholar] [CrossRef]
Peacock, TP; Goldhill, DH; Zhou, J .; Baillon, L .; Frize, R .; Swann, OC; Kugathasan, R .; Penn, R .; Brown, JC; Sanchez-David, RY; et al. The furin cleavage site of SARS-CoV-2 spike protein is a key determinant for transmission due to enhanced replication in airway cells. bioRxiv2020. [Google Scholar] [CrossRef]
Matrajt, L .; Leung, T. Evaluating the Effectiveness of Social Distancing Interventions to Delay or Flatten the Epidemic Curve of Coronavirus Disease. Emerg Infect. Dis.2020, 26, 1740–1748. [Google Scholar] [CrossRef]
Fazio, RH; Ruisch, BC; Moore, CA; Granados Samayoa, JA; Boggs, ST; Ladanyi, JT Social distancing decreases an individual's likelihood of contracting COVID-19. Proc. Natl. Acad. Sci. USA2021, 118, e2023131118. [Google Scholar] [CrossRef] [PubMed]
Gostin, LO; Wiley, LF Governmental Public Health Powers During the COVID-19 Pandemic: Stay-at-home Orders, Business Closures, and Travel Restrictions. JAMA2020, 323, 2137–2138. [Google Scholar] [CrossRef]
Drechsler, Y .; Alcaraz, A .; Bossong, FJ; Collisson, EW; Diniz, PP Feline coronavirus in multicat environments. Vet. Clin. U.S. Small Anim. Pract.2011, 41, 1133–1169. [Google Scholar] [CrossRef]
Ryan, J .; Mazingisa, AV; Wiysonge, CS Cochrane corner: Effectiveness of quarantine in reducing the spread of COVID-19. Mr. Afr. Med. J.2020, 35, 18. [Google Scholar] [CrossRef] [PubMed]
Brooks, SK; Webster, RK; Smith, LE; Woodland, L .; Wessely, S .; Greenberg, N .; Rubin, GJ The psychological impact of quarantine and how to reduce it: Rapid review of the evidence. Lancet2020, 395, 912–920. [Google Scholar] [CrossRef]
Scott, FW Evaluation of risks and benefits associated with vaccination against coronavirus infections in cats. Adv. Vet. Med.1999, 41, 347–358. [Google Scholar] [CrossRef] [PubMed]
Fehr, D .; Holznagel, E .; Bolla, S .; Hauser, B .; Herrewegh, AA; Horzinek, MC; Lutz, H. Placebo-controlled evaluation of a modified life virus vaccine against feline infectious peritonitis: Safety and efficacy under field conditions. Vaccine1997, 15, 1101–1109. [Google Scholar] [CrossRef]
Balint, A .; Farsang, A .; Szeredi, L .; Zadori, Z .; Belak, S. Recombinant feline coronaviruses as vaccine candidates confer protection in SPF but not in conventional cats. Vet. Microbiol.2014, 169, 154–162. [Google Scholar] [CrossRef]
Vennema, H .; de Groot, RJ; Harbor, DA; Dalderup, M .; Gruffydd-Jones, T .; Horzinek, MC; Spaan, WJ Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization. J. Virol.1990, 64, 1407–1409. [Google Scholar] [CrossRef] [PubMed]
Polack, FP; Thomas, SJ; Kitchin, N .; Absalon, J .; Gurtman, A .; Lockhart, S .; Perez, JL; Perez Marc, G .; Moreira, ED; Zerbini, C .; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med.2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
Baden, LR; El Sahly, HM; Essink, B .; Kotloff, K .; Frey, S .; Novak, R .; Diemert, D .; Spector, SA; Rouphael, N .; Creech, CB; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med.2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
Sadoff, J .; Gray, G .; Vandebosch, A .; Cardenas, V .; Shukarev, G .; Grinsztejn, B .; Goepfert, PA; Truyers, C .; Fennema, H .; Spiessens, B .; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. N. Engl. J. Med.2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
Voysey, M .; Clemens, SAC; Madhi, SA; Weckx, LY; Folegatti, PM; Aley, PK; Angus, B .; Baillie, VL; Barnabas, SL; Bhorat, QE; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomized controlled trials in Brazil, South Africa, and the UK. Lancet2021, 397, 99–111. [Google Scholar] [CrossRef]
Tanriover, MD; Doganay, HL; Akova, M .; Guner, HR; Azap, A .; Akhan, S .; Kose, S .; Erdinc, FS; Akalin, EH; Tobacco, OF; et al. Efficacy and safety of an inactivated whole-virion SARS-CoV-2 vaccine (CoronaVac): Interim results of a double-blind, randomized, placebo-controlled, phase 3 trial in Turkey. Lancet2021, 398, 213–222. [Google Scholar] [CrossRef]
Ella, R .; Vadrevu, KM; Jogdand, H .; Prasad, S .; Reddy, S .; Sarangi, V .; Ganneru, B .; Sapkal, G .; Yadav, P .; Abraham, P .; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBV152: A double-blind, randomized, phase 1 trial. Lancet Infect. Dis.2021, 21, 637–646. [Google Scholar] [CrossRef]
Ali Waggiallah, H. Thrombosis Formation after COVID-19 Vaccination Immunological Aspects: Review Article. Saudi J. Biol. Sci.2021, 29, 1073–1078. [Google Scholar] [CrossRef]
Schultz, NH; Sorvoll, IH; Michelsen, AE; Munthe, LA; Lund-Johansen, F .; Ahlen, MT; Wiedmann, M .; Aamodt, AH; Skattor, TH; Tjonnfjord, GE; et al. Thrombosis and Thrombocytopenia after ChAdOx1 nCoV-19 Vaccination. N. Engl. J. Med.2021, 384, 2124–2130. [Google Scholar] [CrossRef]
See, I .; Lale, A .; Marquez, P .; Streiff, MB; Wheeler, AP; Tepper, NK; Woo, EJ; Broder, KR; Edwards, KM; Gallego, R .; et al. Case Series of Thrombosis with Thrombocytopenia Syndrome after COVID-19 Vaccination-United States, December 2020 to August 2021. Ann. Intern. Med.2022. [Google Scholar] [CrossRef]
Dagan, N .; Barda, N .; Kepten, E .; Miron, O .; Perchik, S .; Katz, MA; Hernan, MA; Lipsitch, M .; Reis, B .; Balicer, RD BNT162b2 mRNA Covid-19 Vaccine in a Nationwide Mass Vaccination Setting. N. Engl. J. Med.2021, 384, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
Hall, VJ; Foulkes, S .; Saei, A .; Andrews, N .; Oguti, B .; Charlett, A .; Wellington, E .; Stowe, J .; Gillson, N .; Atti, A .; et al. COVID-19 vaccine coverage in health-care workers in England and effectiveness of BNT162b2 mRNA vaccine against infection (SIREN): A prospective, multicenter, cohort study. Lancet2021, 397, 1725–1735. [Google Scholar] [CrossRef]
Pawlowski, C .; Lenehan, P .; Puranik, A .; Agarwal, V .; Venkatakrishnan, AJ; Niesen, MJM; O'Horo, JC; Virk, A .; Swift, MD; Badley, AD; et al. FDA-authorized mRNA COVID-19 vaccines are effective for real-world evidence synthesized across a multi-state health system. Med2021, 2, 979–992. [Google Scholar] [CrossRef] [PubMed]
Levine-Tiefenbrun, M .; Yelin, I .; Katz, R .; Herzel, E .; Golan, Z .; Schreiber, L .; Wolf, T .; Nadler, V .; Ben-Tov, A .; Kuint, J .; et al. Initial report of decreased SARS-CoV-2 viral load after inoculation with the BNT162b2 vaccine. Nat. Med.2021, 27, 790–792. [Google Scholar] [CrossRef] [PubMed]
Holzworth, J. Infectious diseases of cats. Cornell Vet.1963, 53, 131–143. [Google Scholar]
Weiss, RC; Oostrom-Ram, T. Inhibitory effects of ribavirin alone or combined with human alpha interferon on feline infectious peritonitis virus replication in vitro. Vet. Microbiol.1989, 20, 255–265. [Google Scholar] [CrossRef]
Weiss, RC; Cox, NR; Martinez, ML Evaluation of free or liposome-encapsulated ribavirin for antiviral therapy of experimentally induced feline infectious peritonitis. Res. Vet. Sci.1993, 55, 162–172. [Google Scholar] [CrossRef]
Khalili, JS; Zhu, H.; Mak, NSA; Yan, Y .; Zhu, Y. Novel coronavirus treatment with ribavirin: Groundwork for an evaluation concerning COVID-19. J. Med. Virol.2020, 92, 740–746. [Google Scholar] [CrossRef]
Zeng, YM; Xu, XL; He, XQ; Tang, SQ; Li, Y .; Huang, YQ; Harypursat, V .; Chen, YK Comparative effectiveness and safety of ribavirin plus interferon-alpha, lopinavir / ritonavir plus interferon-alpha, and ribavirin plus lopinavir / ritonavir plus interferon-alpha in patients with mild to moderate novel coronavirus disease 2019: Study protocol. Chin. Med. J.2020, 133, 1132–1134. [Google Scholar] [CrossRef]
Dickinson, PJ; Bannasch, M .; Thomasy, SM; Murthy, VD; Vernau, KM; Liepnieks, M .; Montgomery, E .; Knickelbein, KE; Murphy, B .; Pedersen, NC Antiviral treatment using the adenosine nucleoside analogue GS-441524 in cats with clinically diagnosed neurological feline infectious peritonitis. J. Vet. Intern. Med.2020, 34, 1587–1593. [Google Scholar] [CrossRef]
Pedersen, NC; Perron, M .; Bannasch, M .; Montgomery, E .; Murakami, E .; Liepnieks, M .; Liu, H. Efficacy and safety of the nucleoside analog GS-441524 for the treatment of cats with naturally occurring feline infectious peritonitis. J. Feline Med. Surg.2019, 21, 271–281. [Google Scholar] [CrossRef]
Pedersen, NC; Kim, Y .; Liu, H .; Galasiti Kankanamalage, AC; Eckstrand, C .; Groutas, WC; Bannasch, M .; Meadows, JM; Chang, KO Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. J. Feline Med. Surg.2018, 20, 378–392. [Google Scholar] [CrossRef]
Krentz, D .; Zenger, K .; Alberer, M .; Felten, S .; Bergmann, M .; Dorsch, R .; Matiasek, K .; Kolberg, L .; Hofmann-Lehmann, R .; Meli, ML; et al. Curing Cats with Feline Infectious Peritonitis with an Oral Multi-Component Drug Containing GS-441524. Viruses2021, 13, 2228. [Google Scholar] [CrossRef] [PubMed]
Hartmann, K .; Ritz, S. Treatment of cats with feline infectious peritonitis. Vet. Immunol. Immunopathol.2008, 123, 172–175. [Google Scholar] [CrossRef]
Veronese, N .; Demurtas, J .; Yang, L .; Tonelli, R .; Barbagallo, M .; Lopalco, P .; Lagolio, E .; Celotto, S .; Pizzol, D .; Zou, L .; et al. Use of Corticosteroids in Coronavirus Disease 2019 Pneumonia: A Systematic Review of the Literature. Front. Med.2020, 7, 170. [Google Scholar] [CrossRef] [PubMed]
The WHO Rapid Evidence Appraisal for COVID-19 Therapies (REACT) Working Group. Association Between Administration of Systemic Corticosteroids and Mortality Among Critically Ill Patients With COVID-19: A Meta-analysis. JAMA2020, 324, 1330–1341. [Google Scholar] [CrossRef]
Group, RC; Horby, P .; Lim, WS; Emberson, JR .; Mafham, M .; Bell, JL; Linsell, L .; Staplin, N .; Brightling, C .; Ustianowski, A .; et al. Dexamethasone in Hospitalized Patients with Covid-19. N. Engl. J. Med.2021, 384, 693–704. [Google Scholar] [CrossRef]
Tanaka, Y .; Sato, Y .; Osawa, S .; Inoue, M .; Tanaka, S .; Sasaki, T. Suppression of feline coronavirus replication in vitro by cyclosporin A. Vet. Res.2012, 43, 41. [Google Scholar] [CrossRef]
Tanaka, Y .; Sato, Y .; Takahashi, D .; Matsumoto, H .; Sasaki, T. Treatment of a case of feline infectious peritonitis with cyclosporin A. Vet. Re. Case Rep.2015, 3, e000134. [Google Scholar] [CrossRef]
Cour, M .; Ovize, M .; Argaud, L. Cyclosporine A: A valid candidate to treat COVID-19 patients with acute respiratory failure? Crit. Care2020, 24, 276. [Google Scholar] [CrossRef]
Rudnicka, L .; Glowacka, P .; Goldust, M .; Sikora, M .; Sar-Pomian, M .; Rakowska, A .; Samochocki, Z .; Olszewska, M. Cyclosporine therapy during the COVID-19 pandemic. J. Am. Acad. Dermatol.2020, 83, e151 – e152. [Google Scholar] [CrossRef]
Sanchez-Pernaute, O .; Romero-Bueno, FI; Selva-O'Callaghan, A. Why choose cyclosporin A as first-line therapy in COVID-19 pneumonia. Rheumatol. Clin.2021, 17, 555–557. [Google Scholar] [CrossRef] [PubMed]
Softic, L .; Brillet, R .; Berry, F .; Ahnou, N .; Nevers, Q .; Morin-Dewaele, M .; Hamadat, S .; Bruscella, P .; Fourati, S .; Pawlotsky, JM; et al. Inhibition of SARS-CoV-2 Infection by the Cyclophilin Inhibitor Alisporivir (Debio 025). Antimicrob. Agents Chemother.2020, 64, e00876-20. [Google Scholar] [CrossRef] [PubMed]
Carbajo-Lozoya, J .; Ma-Lauer, Y .; Malesevic, M .; Theuerkorn, M .; Kahlert, V .; Prell, E .; von Brunn, B .; Muth, D .; Baumert, TF; Drosten, C .; et al. Human coronavirus NL63 replication is cyclophilin A-dependent and inhibited by non-immunosuppressive cyclosporine A-derivatives including Alisporivir. Virus Res.2014, 184, 44–53. [Google Scholar] [CrossRef]
Hugo, TB; Heading, KL Prolonged survival of a cat diagnosed with feline infectious peritonitis by immunohistochemistry. Can. Vet. J.2015, 56, 53–58. [Google Scholar] [PubMed]
Conforti, C .; Giuffrida, R .; Zalaudek, I .; Di Meo, N. Doxycycline, a widely used antibiotic in dermatology with a possible anti-inflammatory action against IL-6 in COVID-19 outbreak. Dermatol. Ther.2020, 33, e13437. [Google Scholar] [CrossRef] [PubMed]
Izes, AM; Yu, J .; Norris, JM; Govendir, M. Current status on treatment options for feline infectious peritonitis and SARS-CoV-2 positive cats. Vet. Q.2020, 40, 322–330. [Google Scholar] [CrossRef] [PubMed]
Hung, IF; Lung, KC; Tso, EY; Liu, R .; Chung, TW; Chu, MY; Ng, YY; Lo, J .; Chan, J .; Tam, AR; et al. Triple combination of interferon beta-1b, lopinavir-ritonavir, and ribavirin in the treatment of patients admitted to hospital with COVID-19: An open-label, randomized, phase 2 trial. Lancet2020, 395, 1695–1704. [Google Scholar] [CrossRef]
Doki, T .; Takano, T .; Kawagoe, K .; Kito, A .; Hohdatsu, T. Therapeutic effect of anti-feline TNF-alpha monoclonal antibody for feline infectious peritonitis. Res. Vet. Sci.2016, 104, 17–23. [Google Scholar] [CrossRef] [PubMed]
Luo, P .; Liu, Y .; Qiu, L .; Liu, X .; Liu, D .; Li, J. Tocilizumab treatment in COVID-19: A single center experience. J. Med. Virol.2020, 92, 814–818. [Google Scholar] [CrossRef]
Capra, R .; De Rossi, N .; Mattioli, F .; Romanelli, G .; Scarpazza, C .; Sormani, MP; Cossi, S. Impact of low dose tocilizumab on mortality rate in patients with COVID-19 related pneumonia. Eur. J. Intern. Med.2020, 76, 31–35. [Google Scholar] [CrossRef]
Radbel, J .; Narayanan, N .; Bhatt, PJ Use of Tocilizumab for COVID-19-Induced Cytokine Release Syndrome: A Cautionary Case Report. Chest2020, 158, e15 – e19. [Google Scholar] [CrossRef] [PubMed]
Pediatric Intensive Care Society Statement: Increased Number of Reported Cases of Novel Presentation of Multisystem Inflammatory Disease; Pediatric Intensive Care Society: London, UK, 2020.
Dufort, EM; Koumans, EH; Chow, EJ; Rosenthal, EM; Muse, A .; Rowlands, J .; Barranco, MA; Maxted, AM; Rosenberg, ES; Easton, D .; et al. Multisystem Inflammatory Syndrome in Children in New York State. N. Engl. J. Med.2020, 383, 347–358. [Google Scholar] [CrossRef] [PubMed]
Feldstein, LR; Tenforde, MW; Friedman, KG; Newhams, M .; Rose, EB; Dapul, H .; Soma, VL; Maddux, AB; Mourani, PM; Bowens, C .; et al. Characteristics and Outcomes of US Children and Adolescents With Multisystem Inflammatory Syndrome in Children (MIS-C) Compared With Severe Acute COVID-19. JAMA2021, 325, 1074–1087. [Google Scholar] [CrossRef]
Tullie, L .; Ford, K .; Bisharat, M .; Watson, T .; Thakkar, H .; Mullassery, D .; Giuliani, S .; Blackburn, S .; Cross, K .; De Coppi, P .; et al. Gastrointestinal features in children with COVID-19: An observation of varied presentation in eight children. Lancet Child Adolesc. Health2020, 4, e19 – e20. [Google Scholar] [CrossRef]
LaRovere, KL; Riggs, BJ; Poussaint, TY; Young, CC; Newhams, MM; Maamari, M .; Walker, TC; Singh, AR; Dapul, H .; Hobbs, CV; et al. Neurologic Involvement in Children and Adolescents Hospitalized in the United States for COVID-19 or Multisystem Inflammatory Syndrome. JAMA Neurol.2021, 78, 536–547. [Google Scholar] [CrossRef] [PubMed]
Feldstein, LR; Rose, EB; Horwitz, SM; Collins, JP; Newhams, MM; Son, MBF; Newburger, JW; Kleinman, LC; Heidemann, SM; Martin, AA; et al. Multisystem Inflammatory Syndrome in US Children and Adolescents. N. Engl. J. Med.2020, 383, 334–346. [Google Scholar] [CrossRef]
Blumfield, E .; Levin, TL; Kurian, J .; Lee, EY; Liszewski, MC Imaging Findings in Multisystem Inflammatory Syndrome in Children (MIS-C) Associated with Coronavirus Disease (COVID-19). AJR Am. J. Roentgenol.2021, 216, 507–517. [Google Scholar] [CrossRef] [PubMed]
Alberer, M .; von Both, U. Cats and kids: How a feline disease may help us unravel COVID-19 associated paediatric hyperinflammatory syndrome. Infection2021, 49, 191–193. [Google Scholar] [CrossRef]
Sharma, C .; Ganigara, M .; Galeotti, C .; Burns, J .; Berganza, FM; Hayes, DA; Singh-Grewal, D .; Bharath, S .; Sajjan, S .; Bayry, J. Multisystem inflammatory syndrome in children and Kawasaki disease: A critical comparison. Nat. Rev. Rheumatol.2021, 17, 731–748. [Google Scholar] [CrossRef]
Groff, D .; Sun, A .; Ssentongo, AE; Ba, DM; Parsons, N .; Poudel, GR; Lekoubou, A .; Oh, JS; Ericson, JE; Ssentongo, P .; et al. Short-term and Long-term Rates of Postacute Sequelae of SARS-CoV-2 Infection: A Systematic Review. JAMA Netw. Open2021, 4, e2128568. [Google Scholar] [CrossRef] [PubMed]
McAloose, D .; Laverack, M .; Wang, L .; Killian, ML; Caserta, LC; Yuan, F .; Mitchell, PK; Queen, K .; Mauldin, MR; Cronk, BD; et al. From People to Panthera: Natural SARS-CoV-2 Infection in Tigers and Lions at the Bronx Zoo. mBio2020, 11, e02220-20. [Google Scholar] [CrossRef] [PubMed]
Gaudreault, NN; Trujillo, JD; Carossino, M .; Meekins, DA; Morozov, I .; Madden, DW; Indran, SV; Bold, D .; Balaraman, V .; Kwon, T .; et al. SARS-CoV-2 infection, disease and transmission in domestic cats. Emerg Microbes Infect2020, 9, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
Miro, G .; Regidor-Cerrillo, J .; Czech Republic, R .; Diezma-Diaz, C .; Montoya, A .; Garcia-Cantalejo, J .; Botias, P .; Arroyo, J .; Ortega-Mora, LM SARS-CoV-2 Infection in One Cat and Three Dogs Living in COVID-19-Positive Households in Madrid, Spain. Front. Vet. Sci.2021, 8, 779341. [Google Scholar] [CrossRef] [PubMed]
Giraldo-Ramirez, S .; Rendon-Marin, S .; Jaimes, JA; Martinez-Gutierrez, M .; Ruiz-Saenz, J. SARS-CoV-2 Clinical Outcome in Domestic and Wild Cats: A Systematic Review. Animals2021, 11, 2056. [Google Scholar] [CrossRef] [PubMed]
Ferasin, L .; Fritz, M .; Ferasin, H .; Becquart, P .; Corbet, S .; Ar Gouilh, M .; Legros, V .; Leroy, EM Infection with SARS-CoV-2 variant B.1.1.7 detected in a group of dogs and cats with suspected myocarditis. Vet. Rec.2021, 189, e944. [Google Scholar] [CrossRef] [PubMed]
Chiba, S .; Halfmann, PJ; Hatta, M .; Maemura, T .; Fan, S .; Armbrust, T .; Swartley, OM; Crawford, LK; Kawaoka, Y. Protective Immunity and Persistent Lung Sequelae in Domestic Cats after SARS-CoV-2 Infection. Emerg. Infect. Dis.2021, 27, 660–663. [Google Scholar] [CrossRef]
This article discusses the development of knowledge about feline infectious peritonitis (FIP) from its recognition in 1963 to the present and has been prepared to inform veterinarians, cat rescuers and carers, shelter staff and cat lovers. The causative agent of the feline coronavirus and its relationship to the ubiquitous and minimally pathogenic feline intestinal coronavirus, epizootology, pathogenesis, pathology, clinical signs and diagnostics are briefly mentioned. The main emphasis is placed on the risk factors influencing the incidence of FIP and the role of modern antivirals in successful treatment.
Introduction
Figure 1. Photo of the author and Dr. Jean Holzworthová (1915-2007) from 1991. Dr. Holzworth was the best feline veterinarian the author knew and was responsible for the first report of FIP as a specific disease. She spent her entire career at Angell Memorial Animal Hospital in Boston.
Feline infectious peritonitis (FIP) was described as a specific disease in 1963 by veterinarians at Angell Memorial Animal Hospital in Boston (Holzworth 1963) (Fig. 1). Pathology records from this institution and Ohio State University failed to identify earlier cases (Wolfe and Griesemer 1966), although identical cases were soon recognized worldwide. The initial pathological descriptions were of diffuse inflammation of the tissues lining the abdominal cavity and abdominal organs with extensive effusion of inflammatory fluid, after which the disease was eventually named (Wolfe and Griesemer 1966, 1971) (Figs. 2, 3). A second and less common clinical form of FIP, which presents with less diffuse and more widespread granulomatous lesions involving organ parenchyma, was first described in 1972 (Montali and Strandberg 1972) (Figs. 3,4). The presence of inflammatory effusions in the body cavity in the common form and the absence of effusions in the less common form led to the names wet (effusion, non-parenchymatous) and dry (non-effusion, parenchymatous) FIP.
The prevalence of FIP appears to have increased during the panzootic disease caused by feline leukemia virus (FeLV) in the 1960s–1980s, when many cases of FIP were found to be associated with FeLV (Cotter et al., 1973; Pedersen 1976a). The subsequent management of FeLV infection in domestic cats through rapid testing and vaccination resulted in an increase in the number of FIP cases. However, recent interest in breeding/rescue along with effective treatment has led to increased awareness of the disease and its diagnosis.
Figure 2. Gross necroptic appearance of the abdominal cavity of a cat with acute onset wet FIP. The abdomen is filled with several hundred ml of yellow viscous fluid, the omentum is reddened, edematous and contracted, and fibrin deposits (arrows) are visible on the surface of the spleen and the edges of the liver. A fiber of fibrin can be seen on the spleen
Figure 3. Appearance of an open abdomen at autopsy of a cat that died of a chronic form of effusive FIP. The abdomen is filled with a viscous, yellow-colored exudate, and the omentum is thickened and contracted. The main lesions are in the liver with numerous plaque-like structures (pyogranulomas) on the cover. More circumscribed lesions (granulomas), also oriented on the serous surface, look more fleshy and are elevated above the surface. These lesions also involve the underlying liver parenchyma and are more typical of dry FIP. This is an example of a case of FIP that transitions between the wet and dry forms (arrow).
Figure 4A – Gross section of kidneys of two cats with dry form of FIP. The lesions are superficial and extend into the underlying parenchyma.
Figure 4B - lesions of the dry form of FIP in organs such as the kidneys, cecum, colon and intestinal lymph nodes (Fig. 5) were grossly confused with renal lymphoma.
Figure 5. Gross enlargement of ileo-cecal-colic lymph nodes in a cat with dry FIP.
Etiological factor
The first attempts did not allow identifying the causative agent of FIP, but confirmed its infectious nature (Wolfe and Griesemer 1966). A viral etiology was established in 1968 using ultrafiltrates of infectious material (Zook et al., 1968). The causative virus was subsequently identified as a coronavirus (Ward 1970), which is closely related to enteric coronaviruses of dogs and pigs (Pedersen et al., 1978).
Confusion arose when feline enteric coronavirus (FECV) was isolated from the feces of healthy cats and proved to be indistinguishable from feline infectious peritonitis virus (FIPV) (Pedersen et al., 1981). Unlike FIPV, which readily induced FIP in laboratory cats, experimental infections with FECV were largely asymptomatic. The relationship between the two viruses became clear when FIPVs were found to be FECV mutants that arise in the body of every cat with FIP (Vennema et al., 1995; Poland et al., 1996).
FECV is ubiquitous in feline populations worldwide and is first shed in faeces from approximately 9–10 weeks of age, coinciding with the loss of maternal immunity (Pedersen et al., 2008 ;). The infection takes place via the faecal-oral route and targets the intestinal epithelium, and the primary signs of enteritis are mild or inconspicuous (Pedersen et al., 2008; Vogel et al., 2010). Subsequent faecal excretion occurs from the colon and usually stops after several weeks or months (Herrewegh et al., 1997; Pedersen et al., 2008; Vogel et al., 2010). Immunity is short-lived and repeated infections are common (Pedersen et al., 2008; Pearson et al., 2016). Over time, stronger immunity eventually develops and cats older than 3 years are less likely to shed the infection in their faeces (Addie et al., 2003). FECV is constantly subject to genetic drift into locally and regionally identifiable clades (Herrewegh et al., 1997; Pedersen et al., 2009).
FECV and FIPV are classified as biotypes of the feline coronavirus (FCoV) subspecies. The genomes of FECV and FIPV biotypes are related at >98 %, but with unique host cell tropism and pathogenicity (Chang et al., 2012; Pedersen et al., 2009). FECVs infect the mature intestinal epithelium, whereas FIPVs lose intestinal tropism and acquire the ability to replicate in monocytes/macrophages. The published names FECV or FIPV will be used here when discussing aspects of the disease specific to each biotype, while the term FCoV will be used when discussing features common to both biotypes.
Three types of mutations are involved in the biotype change of FECV to FIPV. The first type, which is unique to each cat with FIP (Poland et al., 1996), consists of an accumulation of missense and nonsense mutations in the c-terminus of the auxiliary 3c gene, often resulting in truncated 3c gene products (Pedersen et al., 2012 ; Vennema et al., 1995). The second type of mutation consists of two specific single nucleotide polymorphisms in the fusion peptide of the S gene, one or the other form being common to >95 % FIPV and absent in FECV (Chang et al., 2012). A third type of mutation, unique to each FIPV isolate and not found in FECV, occurs in and around the furin cleavage motif between the receptor binding domain (S1) and the fusion domain (S2) of the spike gene (S) (Licitra et al., 2013). These mutations have different effects on furin cleavage activity. Together and in an as yet undetermined manner, they are responsible for the shift of the tropism of the host cell from the enterocyte to the macrophage and for the profound change in the form of the disease.
FCoV, and therefore FECV and FIPV, exist in two serotypes identified by antibodies against the viral neutralizing epitope on the S gene (Herrewegh et al., 1998; Terada et al., 2014). Serotype I FCoVs are identified in cat sera and are prevalent in most countries. Serotype II FCoVs result from recombination with the S part of the canine coronavirus gene (Herrewegh et al., 1998; Terada et al., 2014) and are identified by canine coronavirus antibodies. Serotype II FIPVs are easily cultured in tissue culture, whereas serotype I FIPVs are difficult to adapt to growth in vitro. Serotype I and II FECVs were not grown in conventional cell cultures (Tekes et al., 2020).
FIPVs are found exclusively in activated monocytes and macrophages in affected tissues and effusions and are not secreted into the environment. Therefore, cat-to-cat (horizontal) transmission of FIPV is not the main mode of spread. Rather, FIP follows the pattern of an underlying enzootic FECV infection, with sporadic cases and occasional small outbreaks of disease (Foley et al., 1997). These clusters of cases can be mistaken for epizootics. The only report of an epizootic occurrence of FIP was associated with a single serotype II virus that appeared to develop in a shelter housing both dogs and cats (Wang et al., 2013). Horizontal transmission in this case followed an epizootic rather than an enzootic disease model, with infection spreading rapidly to cats of all ages and in close contact with the index case (Wang et al., 2013).
The low incidence of FIP cases in the population suggests that FIPV mutations arise infrequently. However, studies involving FECV infection in immunocompromised cats infected with FIV and FeLV suggest that FIP mutants may be common but only cause disease under certain circumstances. Nineteen cats infected with feline immunodeficiency virus (FIV) for 6 years and a control group of 20 littermates not infected with FIV were orally challenged with FECV (Poland et al., 1996). Cats in both groups remained asymptomatic for two months when two cats in the FIV-infected group developed FIP. In a second study, 26 young cats with enzootic FECV infection from a breeding colony with no history of FIP were contact-exposed to FeLV carriers (Pedersen et al., 1977). Two kittens in the group subsequently developed FIP 2–10 weeks after becoming FeLV viremic. The question remains, how long can FIPV viruses survive in the body before they are eliminated? According to one of the theories, they persist in the body for a certain time and become pathological only if immunity against them is impaired (Healey et al., 2022). This theory is supported by the way immunity to FeLV develops. Most cats resist FeLV by kitten age and develop robust and permanent immunity, but this occurs within a few weeks, during which the virus persists in a subclinical or latent state (Pedersen et al., 1982; Rojko et al., 1982). . Methylprednisolone given during this period, but not after, will abolish developing immunity and lead to a state of persistent viremia.
Epizootology
Epizootiology is the study of the occurrence, spread and possible control of animal diseases and the influence of environmental, host and agent factors. FIP is considered one of the most important infectious causes of death in cats, although there are no precise data on prevalence. It is estimated that 0.3–1.4 % deaths of cats presented to veterinary institutions are related to FIP (Rohrbach et al., 2001; Pesteanu-Somogyi et al., 2006; Riemer et al., 2016) and in some shelters and breeding stations up to 3.6–7.8 % (Cave et al., 2002). FIP is also described as an environmental disease with a higher incidence of multiple cats. Three-quarters of the FIP cases in the currently ongoing treatment study came from the field through foster carers/rescues and cat shelters, 14 % from kennels, and only 11 % from households.1
Studies based on cases observed in academic institutions have demonstrated the influence of age and gender on the incidence of FIP (Rohrbach et al., 2001; Pesteanu-Somogyi et al., 2006; Pedersen 1976a; Worthing et al., 2012; Riemer et al., 2016) . Three-quarters of the cases in these cohorts occurred in cats younger than 3 years of age, and few cases occurred after 7 years of age. This was also confirmed by a current and ongoing field study from the Czech Republic and Slovakia, in which it was found that more than 80 % cases of FIP occurred in cats under 3 years of age and only 5 % in cats older than 7 years (Fig. 6) .1 Earlier institutional studies differed on the effect of sex, but indications were that male cats were slightly more susceptible to FIP than female cats. This was also confirmed by current data from the field, which show a ratio of males to females of 1.3:1.1. It is unclear whether castration affects the incidence of FIP, with some reports suggesting that it may increase susceptibility (Riemer et al., 2016), while others do not report such a clear effect.1
Figure 6. Age of more than 607 cats from the Czech Republic and Slovakia at the time of diagnosis and treatment of FIP.1 Thirty percent of infections were in cats six months of age or younger, 50 % at one year of age, and 85 % at three years of age or younger.
Other environmental and viral risk factors have been implicated in the increased incidence of FIP, but their significance requires knowledge of disease occurrence in their absence. A possible baseline may have been provided by a study of enzootic FECV infection, which had been unrecognized for many years in a well-managed specific pathogen-free breeding colony (Hickman et al., 1995). This colony was kept in strict quarantine free of other infections and the standard of nutrition and husbandry was high. This colony produced hundreds of kittens each year before the first case of FIP was diagnosed. Such observations suggest that FIP may be a rare phenomenon in the absence of risk factors.
The importance of moving to a new home as a risk factor for FIP is only now being appreciated. Breeders, many of whom have not experienced any cases of FIP in their litters, are most concerned about the announcement that one of their kittens has developed FIP shortly after going to a new home. A recent study found that more than half of cats with FIP had experienced a change in environment, shelter or capture in the weeks before the illness.1 Cats are known to hide external signs of stress even when they are suffering from serious internal ill effects. Even simple procedures such as changing the cage will suppress immunity and reactivate latent herpes virus shedding and disease symptoms in cats (Gaskell and Povey, 1977). Stressful situations, even those that appear to be minor, can cause a decrease in lymphocyte levels and “sick behavior” (Stella et al., 2013).
Differences in the genetic make-up of enzootic FCoV strains may also contribute to the prevalence of FIP in the population. Serotype II FIPVs are thought to be more virulent than serotype I and more likely to be transmitted from cat to cat (Lin et al., 2009; Wang et al., 2013). It is also possible that certain FECV clades are more susceptible to mutation to FIPV, which should be studied. The author also observed a disproportionately high proportion of cats with neurologic FIP in some regions, suggesting that genetic determinants in certain FCoV strains may be more neurotropic.
Immunodeficiencies associated with retroviruses are associated with susceptibility to FIP. Up to half of FIP cases during the peak of FeLV panzootic disease were persistently infected with FeLV (Cotter et al., 1973; Pedersen 1976a; Hardy 1981). FeLV infection causes suppression of T-cell immunity, which may inhibit the protective immune response to FIP. The importance of FeLV infection for the incidence of FIP has declined significantly since the 1980s, when carrier elimination and vaccination pushed FeLV back into the wild, where exposures are less severe and immunity is the usual outcome. Chronic feline immunodeficiency virus (FIV) infection has also been shown to be a risk factor for FIP in FECV-infected cats under experimental conditions (Poland et al., 1996). In one recent field study, FeLV infection was recognized in 2 % and FIV in 1 % cats treated for FIP.1
The incidence of FIP in purebred cats is reported to be higher than in random breeding cats, with some breeds appearing to be more susceptible than others (Pesteanu-Somogyi et al., 2006; Worthing et al., Genetic predisposition to FIP has been investigated in several Persian cat breeds and is estimated to account for half the risk of the disease (Foley et al., 1997). Some breeds, such as the Birman, are more susceptible to developing dry than wet FIP (Golovko et al., 2013). Attempts to identify specific genes associated with susceptibility for FIP in Burmese cats included several immune-related genes, but none reached the desired significance (Golovko et al., 2013).The largest study of genetic susceptibility to FIP showed that it is extremely polymorphic and reported consanguinity as a major risk factor. breeding (Pedersen et al., 2016).Specific polymorphisms in several genes have also been associated with high levels of FECV shedding among several breeding cat breeds (Bubeniko and et al., 2020).
In females, FIP, usually the wet form, may develop during pregnancy or in the perinatal period. This phenomenon resembles the suppression of immunity in pregnant women and the predisposition to certain infections (Mor and Cardenas 2010). It is not clear whether subclinical FIP is activated by pregnancy or by increased susceptibility to new infection. Maternal infection early in pregnancy results in fetal death and resorption, while later infections often result in abortion (Fig. 7). Kittens can be born healthy, but develop disease in the perinatal period and die. Some babies are born uninfected thanks to the effectiveness of the placental barrier between mother and fetus or thanks to the help of antiviral treatment (Fig. 8).
Figure 7. Aborted kittens from a dam that developed wet FIP late in pregnancy. Miscarriage was the first symptom of FIP, quickly followed by the classic symptoms of abdominal wet FIP. The mother was successfully cured of FIP with the antiviral GS-441524.
Figure 8. This mother developed symptoms of wet abdominal FIP 3 weeks after the onset of pregnancy and was successfully treated with GS-441524. Subsequently, she gave birth to a litter of four kittens by caesarean section, one of which died and three survived and grew up healthy. Treatment was given during the remaining 6 weeks of gestation and continued for 6 weeks during which the kittens were successfully nursed. GS-441524 had no apparent side effects on the mother or the kittens.
A possible increase in the number of cases of FIP was observed in cats older than 10 years in studies conducted 50 years ago (Pedersen 1976a). Slightly more than 3 % cases of FIP in a recent study occurred in cats 10 years of age and older and 1.5 % in cats 12 years of age and older (Fig. 6).1 The occurrence of FIP in the elderly often involves two different scenarios. The first scenario also involves exposure to FECV faecal excretion, but in a unique way. It is common for old cats to mate as kittens and live together in relative isolation unexposed to FECV for many years. One cat in the pair dies, is left alone, and a much younger companion obtained from a rescue organization, shelter, or kennel is brought into the household that has a high probability of excreting FECV. Older cats are also susceptible to the same FIP risk factors as younger cats, as well as other factors associated with aging. The first of these is the impact of aging on the immune system, with the most consequential being the deterioration of cellular immune function (Day 2010). Other risk factors associated with old cats include the debilitating and potentially immunosuppressive effects of diseases such as cancer and chronic diseases of the kidneys, liver, oral cavity and intestines. Some diseases in old cats can be mistaken for FIP or complicate the treatment of FIP if they are present at the same time.
Other risk factors that need further investigation include loss of maternal systemic immunity by separation at birth, early weaning and loss of lactogenic immunity, malnutrition, common kitten infectious diseases, early neutering, vaccination, congenital heart defects, and even a shelter fire (Drechsler et al.), 2011; Healey et al., 2022; Pedersen 2009, Pedersen et al. 2019).1 However, the most important positive risk factor remains the presence of FECV in the population (Addie et al., 1995). The prevalence of FIP in several Persian cat breeds was also related in one study to the proportion of cats that shed FECV at a given time and to the proportion of these cats that are chronic shedders (Foley et al., 1997). The importance of exposure to FECV supports the need to find ways to either prevent infection or reduce its severity. One of the first steps is a better understanding of FECV immunity (Pearson et al., 2019).
Pathogenesis
The first interface between FECV and the immune system is the lymphatic tissues of the intestine (Malbon et al., 2019, 2020). Although the downstream events leading to FIP are not fully understood, it is possible to speculate based on what is already known about FECV and FIPV infections, other macrophage-tropic infections, and viral immunity in general. During intestinal infection, FECV particles and proteins reach the local lymphatic tissues and are processed by phagocytic cells first into peptides and finally into amino acids. Some of these peptides will be recognized as foreign when arrayed on the cell surface, triggering innate (innate or non-specific) and adaptive (acquired or specific) immune responses (Pearson et al., 2016). FECVs also mutate to FIPV at the same time and in the same cell type. Some of these mutations will allow the virus to replicate in these or closely related cells of a specific monocyte/macrophage lineage.
The host cell for FIPV appears to be a specific class of activated monocytes found around venules on the surface of intestinal and thoracic organs, mesentery, omentum, uveal tract, meninges, choroid and ependyma of the brain and spinal cord, and freely in effusions. These cells belong to the activated (M1) class (Watanabe et al., 2018) and resemble a subpopulation of small peritoneal macrophages described in mice (Cassado et al., 2015). This type of cell arises from circulating bone marrow-derived monocytes that are rapidly mobilized from the blood in response to infectious or inflammatory stimuli. A similar-looking population of activated monocytes has been described around blood vessels in the retina affected by FIP (Ziolkowska et al., 2017). These cells stained for calprotectin, indicating their blood origin. Although FIPV infection occurs initially in smaller activated monocytes, viral replication is most intense in large, vacuolated, terminally differentiated macrophages (Watanabe et al., 2018). The virus released from these cells rapidly infects activated monocytes produced in the bone marrow and drawn to the site from the bloodstream.
The cellular receptor used by FECVs to infect intestinal epithelial cells has not yet been determined. The cellular receptor that FIPVs use to infect activated monocytes is also unknown. RNAs for conventional coronavirus receptors such as aminopeptidase N (APN), angiotensin converting enzyme 2 (ACE2) and CD209L (L-SIGN) were not upregulated in infected peritoneal cells of cats with experimental FIP, and CD209 (DC-SIGN) was significantly underexpressed (Watanabe et al., 2018). An alternative route of infection of activated monocytes may involve immune complexation of the virus and entry into cells by phagocytosis (Dewerchin et al., 2008, 2014; Van Hamme et al., 2008). Activated monocytes in lesions stain strongly positive for FIPV antigen, IgG and complement (Pedersen, 2009) and mRNA for FcγRIIIA (CD16A/ADCC receptor) is markedly increased in infected cells (Watanabe et al., 2018), supporting infection through immune complexation and alternative receptors related to phagocytosis.
Macrophage pathogens are intracellular and elimination of infected cells occurs through lymphocyte-mediated killing. The first line of defense is non-specific lymphocytes, and if they fail, an adaptive immune response to FIPV follows through specific T-lymphocytes. If infected activated monocytes and macrophages fail to be contained and eliminated, they may disseminate locally in the abdominal cavity, possibly from lymph nodes in the lower intestinal region and the site of FECV replication. Spread locally and to distant sites via the bloodstream is by infected monocyte cells (Kipar et al., 2005).
FIP occurs in two basic forms, wet (effusive, nonparenchymatous) (Figures 2 and 3) or dry (noneffusive, parenchymatous) (Figures 4 and 5), with wet FIP accounting for 80 % cases.1 The term “wet” refers to the characteristic effusion of fluid in the abdomen or chest (Wolfe and Griesemer 1966, 1971). Wet FIP lesions are dominated by inflammation resembling immediate- or Arthus-type hypersensitivity (Pedersen and Boyle, 1980), whereas dry FIP lesions resemble delayed-type hypersensitivity reactions (Montali and Strandberg 1972; Pedersen 2009). Wet and dry forms of FIP therefore reflect competing influences of antibody- and cell-mediated immunity and associated cytokine pathways (Malbon et al., 2020, Pedersen 2009). Immunity to FIPV-infected cells, which is the norm, is thought to involve strong cell-mediated responses (Kamal et al. 2019). Dry FIP is thought to occur when cell-mediated immunity is partially effective in suppressing infection, while wet FIP occurs when cell-mediated immunity is ineffective and humoral immune responses predominate.
FIP is considered unique among macrophage infections because it is viral, but the dry form shares many clinical and pathogenic features with feline diseases caused by systemic mycobacterial (Gunn-Moore et al., 2012) and fungal infections (Lloret et al., 2013). . Similarities in pathogenesis also exist between wet FIP and antibody-enhanced viral infections such as dengue fever and dengue hemorrhagic shock syndrome (Pedersen and Boyle 1980; Rothman et al., 1999; Weiss and Scott 1981).
Host responses are thought to solely determine the outcome of FIPV infection and the resulting forms of disease. However, macrophage-tropic pathogens have evolved their own unique defense mechanisms against the host (Leseigneur et al., 2020). One of the mechanisms is the delay of programmed cell death (apoptosis). Delayed apoptosis allows sustained microbial replication and eventual release of more infectious agents, as has also been described in FIPV-infected macrophages (Watanabe et al., 2018). FIPV can also control the recognition and killing of infected activated monocytes by specific or non-specific T-cells. The cell surface targets for T-cells that kill infected cells are likely FIPV proteins (antigens) expressed on major histocompatibility complex class I (MHC-I) receptors. However, surface expression of viral antigens by MHC-I receptors was not detected on FIPV-positive cells collected from FIP tissues or effusions (Cornelissen et al., 2007). DC-Sign has been proposed as a receptor for FIPV (Regan and Whitaker, 2008), but RNA for DC-Sign is markedly underexpressed by infected peritoneal cells, whereas RNA for Fc (MHC-II) receptors is markedly overexpressed and RNA for MHC -I is reduced (Watanabe et al., 2018). This suggests that the normal mode of infection of host cells may be altered by FIPV to favor infection by phagocytosis instead of binding to specific viral receptors on the cell surface, fusion with the cell membrane, and internalization.
Pathology
Detailed descriptions of the gross and microscopic lesions in the wet form of FIP were first described by Wolfe and Griesemer (1966, 1971). The disease is characterized by vasculitis involving venules in the tissues lining the abdominal or thoracic cavity, organ surfaces, and supporting tissues such as the mesentery, omentum, and mediastinum. The inflammatory process leads to effusions in the abdominal or chest cavity up to a volume of one liter or more (Fig. 2, 3). The underlying lesion is a pyogranuloma, which consists of a focal accumulation of activated monocytic cells in various stages of differentiation, interspersed with non-degenerate neutrophils and sparse numbers of lymphocytes. Pyogranulomas are superficially oriented and appear grossly and microscopically as single and coalescent plaques (Fig. 2).
FIPV antigen is immunohistochemically (IHC) observed only in activated monocytes in lesions and effusions (Litster et al., 2013). Large vacuolated terminally differentiated macrophages are particularly rich in virus (Watanabe et al., 2018), reminiscent of the lepromatous form of leprosy (deSousa et al., 2017). Lymph nodes located near the sites of inflammation are hyperplastic and enlarged.
The relationship between dry and wet FIP was first described in 1972 in a report of cases of unknown etiology with similar pathology (Montali and Strandberg 1972). As the authors state, “this pathological syndrome was characterized by granulomatous inflammation in various organs, but mainly affected the kidneys, visceral lymph nodes, lungs, liver, eyes, and leptomeninges.” Tissue extracts from these lesions induced wet FIP in laboratory cats, confirming that wet and dry FIP are caused by the same agent.
The gross and microscopic pathology of dry FIP resembles that of other macrophage-tropic infections such as feline systemic blastomycosis, histoplasmosis, coccidioidomycosis (Lloret et al., 2013), tuberculosis and leprosy (Gunn-Moore et al., 2012). Lesions of dry FIP mainly involve the abdominal organs (Figs. 5, 6) and are rare in the thoracic cavity (Montali and Strandberg 1972; Pedersen 2009). Lesions are less widespread and focal than in wet FIP, with a tendency to extend from the serous surfaces into the parenchyma of the underlying organs (Figs. 5, 6). The target of the host immune response are small aggregates of infected monocytic cells associated with venules, similar to pyogranulomas in wet FIP, but surrounded by dense accumulations of lymphocytes and plasma cells and variable fibrosis. The florid hyperemia, edema, and microhemorrhage associated with wet FIP are mostly absent, therefore significant effusions in the body cavities are absent. The host response to foci of infection gives the lesions a gross tumor-like appearance (Figs. 5, 6). Infected activated monocytes in the central focus of infection are less dense and contain lower levels of virus than in the wet form (Pedersen 2009;), a feature of the tuberculoid form of leprosy (de Sousa et al., 2017). Lesions in some places, for example on the wall of the large intestine, can cause a dense surrounding zone of fibrosis, which resembles classic tuberculosis granulomas. Transitional forms also exist between wet and dry forms in a small number of cases and are mostly recognizable at autopsy (Fig. 3).
Ocular and neurological FIP are classified as forms of dry FIP (Montali and Strandberg 1972). However, pathology in the uveal tract and retina and in the ependyma and meninges of the brain and spinal cord is intermediate between wet and dry FIP (Fankhauser and Fatzer 1977; Peiffer and Wilcock 1991). This can be explained by the effect of the blood-ocular and blood-brain barrier in protecting these areas from systemic immune reactions.
Clinical characteristics of FIP
The five most common symptoms in cats with FIP, regardless of clinical form and frequency of occurrence, are lethargy, loss of appetite, enlarged abdominal lymph nodes, weight loss, fever, and deteriorating coat.1 These symptoms can appear quickly, within a week, or they can exist for many weeks or even months before a diagnosis is made. The course of the disease tends to be more rapid in cats with wet FIP than with dry FIP, and growth retardation is common in young cats, especially those with more chronic disease. 20 % cats with fever as the main symptom are eventually diagnosed with FIP (Spencer et al., 2017).
The wet form of FIP occurs in approximately 80 % cases, more often in younger cats, and tends to be more severe and more rapidly progressive than the dry form. Abdominal effusion (ascites) is four times more common than pleural effusion, with abdominal distension (Fig. 9) and dyspnea being common symptoms. Pyrexia and jaundice are more common symptoms in cats with wet than dry FIP (Tasker, 2018).
Figure 9. Adult longhair cat with chronic abdominal wet FIP. The cat was in acceptable health except for mild weight loss, lethargy, poor coat quality, and occasional low-grade fever. Abdominal distension was not noted for some time, and the abdominal fluid contained relatively low protein and white blood cell counts.
Figure 9. A young cat who presented with rapid onset of high fever, loss of appetite, abdominal distension and abdominal fluid with a high protein and white blood cell content.
Most cats with dry FIP present with disease symptoms limited to the abdomen and/or chest. The most common clinical signs of dry FIP are palpable or ultrasound-identifiable masses in the kidney (Fig. 4), cecum, colon, liver, and associated lymph nodes (Fig. 5). Lesions of dry FIP usually spare the thoracic cavity and rarely occur in the skin, nasal passages, pericardium, and testes as part of a wider systemic disease.
Neurological and ocular disease are the sole or secondary features of 10 % of all FIP cases and are 10 times more often associated with dry than wet FIP (Pedersen 2009). The neurological and ocular forms of FIP have been classified as forms of dry FIP, but it may be more appropriate to classify them as distinct forms of FIP resulting from the modifying effects of the blood-ocular and blood-brain barriers behind which they occur. These barriers have a strong impact on the nature of eye and central nervous system (CNS) disease and response to antiviral therapy.
Clinical signs of neurologic FIP involve both the brain and spinal cord and include posterior weakness and ataxia, generalized incoordination, seizures, mental dullness, anisocoria, and varying degrees of fecal and/or urinary incontinence (Foley et al., 1998; Dickinson et al., 2020) ( Fig. 10). Extreme intracranial pressure can lead to sudden herniation of the cerebellum and brainstem into the spinal canal and spinal shock syndrome. Prodromal symptoms include compulsive wall or floor licking, litter eating, involuntary muscle twitching, and reluctance or inability to jump to high places. Eye involvement may precede or accompany neurological disease. Neurological FIP is a common phenomenon with antiviral therapy, either occurring during treatment of non-CNS forms of FIP or as a manifestation of disease relapse after treatment cessation (Pedersen et al., 2018, 2019; Dickinson et al., 2020).
Figure 10. A young cat with dry FIP and neurological impairment. The cat is lethargic, emaciated and with poor fur. The fur in the perineal area is wet and stained from urinary incontinence.
Figure 11. This cat's right iris staining was the first sign of FIP-associated uveitis. There is a slight haze in the anterior chamber, and there are fibrin deposits rich in red blood cells on the inside of the cornea. The pupils are also unequal (anisocoria).
Figure 12. A young cat with ocular FIP presenting as anterior uveitis in the right eye with secondary glaucoma causing globe enlargement. The iris has changed color due to inflammation, the vessels at the base of the iris are congested, and there is turbidity of aqueous humor and inflammatory products on the back of the cornea. Intraocular pressure is usually low in uncomplicated uveitis but elevated in cats with glaucoma.
Figure 13. This young cat had anterior uveitis, but her FIP therapy with GS-441524 was delayed, allowing glaucoma to develop in both eyes. Treatment cleared the underlying uveitis and greatly improved external health, but secondary glaucoma and blindness persisted.
Eye involvement is usually obvious and is confirmed by ophthalmoscopic examination of the anterior and posterior chambers. Ocular FIP affects the iris, ciliary bodies, retina, and optic disc to varying degrees (Peiffer and Wilcock, 1991; Ziółkowska et al., 2017; Andrew, 2000). The earliest symptom is often a unilateral change in the color of the iris (Fig. 11). The anterior chamber may appear cloudy and may show high protein levels and water turbidity on refraction. Inflammatory products in the form of activated macrophages, red blood cells, fibrin markers and small blood clots are washed into the anterior chamber. This material often adheres to the back of the cornea as keratic precipitates (Fig. 12). The disease can also affect the retina in tapetal and non-tapetal areas and lead to retinal detachment. Intraocular pressure is usually low, except in cases complicated by involvement of the ciliary body and glaucoma (Fig. 12, 13).
FIP Diagnosis
Signaling, environmental history, clinical signs, and physical examination findings often point to FIP (Tasker, 2018). A thorough physical examination should include body weight and temperature, coat and body condition, manual palpation of the abdomen and abdominal organs, gross assessment of cardiac and pulmonary function, and a cursory examination of the eyes and neurological system. Strong suspicion of an effusion in the abdominal or thoracic cavity may warrant confirmatory aspiration and even in-house fluid analysis as part of the initial examination.
Abnormalities in the complete blood count (CBC) and basic serum biochemical panel are important factors in the diagnosis of FIP (Tasker, 2018; Felten and Hartmann, 2019) and monitoring of antiviral therapy (Pedersen et al., 2018, 2019; Jones et al., 2021). ; Krentz et al., 2021) (Fig. 14). Total leukocyte counts are most likely high in cats with wet FIP, but low counts can occur with severe inflammation. A high leukocyte count is often associated with neutrophilia, lymphopenia, and eosinopenia. Mild to moderate non-regenerative anemia is also frequently seen in both wet and dry FIP. Total protein is usually elevated due to elevated globulin levels, while albumin values tend to be low (Fig. 14). This results in an A:G ratio that is often lower than 0.5-0.6 and is considered one of the most consistent indicators of FIP. However, a low A:G ratio can occur in situations where both albumin and globulin are within the reference range or in other diseases. Therefore, the A:G ratio should not be the only FIP indicator and should always be evaluated in the context of other FIP indicators (Tasker, 2018; Felten and Hartmann, 2019). Serum protein values obtained from most serum chemistry panels are usually adequate. Serum protein electrophoresis can provide additional information, especially if protein values from serum chemistry are questionable (Stranieri et al., 2017).
Figure 14. Complete blood count (CBC) (a) of a young cat with acute wet abdominal FIP. Although the leukocyte count was not elevated, relative but not absolute neutrophilia, relative and absolute lymphopenia, relative and absolute eosinopenia, and unresponsive anemia were noted, indicated by low red blood cells, hematocrit, and hemoglobin with a normal reticulocyte count.
Figure 14. Serum biochemical examination (b) of a young cat with acute wet abdominal FIP. Relevant values in the serum chemistry panel were elevated total protein, low albumin, high globulin, low albumin/globulin (A:G) ratio, and elevated total and direct bilirubin. Liver enzymes were normal except for mildly elevated AST and BUN and creatinine were normal, indicating the absence of significant liver or kidney disease. Globulin values are not always given, but a reasonable estimate can be calculated by subtracting the albumin level from the total protein.
Overreliance on CBC and serum biochemistry abnormalities can lead to diagnostic uncertainty when absent, despite the fact that no test value is consistently abnormal in all cases of FIP (Tasker, 2018)1. The biggest differences are between the clinical form of the disease, with leukocytosis and lymphopenia being more common in cats with wet than with dry FIP (Riemer et al., 2016). Hyperbilirubinemia is common in cats with FIP, but especially in cats with wet FIP (Tasker, 2018). The author also found that many cats with primary neurological FIP show minor or no blood abnormalities. Blood test values for FIP also vary from study to study (Tasker, 2018).
A complete analysis of the effusion is important to diagnose wet FIP and to rule out other potential causes of fluid accumulation (Dempsey and Ewing, 2011). It includes color (clear or yellow), viscosity (thin or viscous), presence of precipitates, ability to form a partial clot on standing, protein content, leukocyte count, and differential. The nature of the fluid may vary depending on the duration of the disease and its severity. Effusions in cats with more severe disease usually have protein values close to serum values, are more viscous, contain more leukocytes, are more yellow in color, and have a greater ability to form partial clots on standing. Chronic effusions tend to be less inflammatory in nature, with lower protein and leukocyte counts, less viscous and clearer. These values can be determined on the spot in most clinics. The clotting factor is determined by comparing the fluid collected in the serum and in the anticoagulant tubes after standing. Color and viscosity can be approximated and protein levels can be estimated using a handheld refractometer to determine total solids. Cells are pelleted from the fluid and analyzed on a fast-stained slide using light microscopy, and the leukocyte count and differential are estimated. Cells include nonseptic neutrophils, small and medium-sized mononuclear cells, and large vacuolated macrophages (Fig. 15). It is important to note that effusions can occur in a variety of conditions, such as heart failure, cancer, hypoproteinemia, and bacterial infections. Effusions in these other diseases usually have different identifying features.
Figure 15. Stained smear of peritoneal cells centrifuged from the abdominal fluid of a cat with wet FIP and examined on a fast-stained slide by light microscopy. The predominant cells are large strongly vacuolated macrophages, smaller differentiated activated monocytes and neutrophils. The greatest concentration of viral particles is in the intracytoplasmic vacuoles of macrophages (arrows).
Figure 16. Positive result of the Rivalt test. A small sample of abdominal or thoracic fluid is carefully dropped into a small beaker filled with diluted acetic acid (8 ml of distilled water and 1 drop of concentrated acetic acid). Inflammatory proteins almost immediately precipitate and sink to the bottom (positive). Less inflammatory fluids will form diffuse precipitates (questionable) or diffuse freely in solution (negative).
A positive Rivalt test on abdominal or chest fluid is often used to diagnose FIP as a cause of effusion, and a negative test tends to rule it out (Fischer et al., 2010) (Fig. 16). However, the test may be positive in inflammatory effusions of another cause and negative in some cats with FIP. Therefore, Rivalt's test is most helpful in combination with other clinical findings of FIP and should not replace a thorough fluid analysis (Felten and Hartmann, 2019).
Serum total and direct bilirubin levels are often elevated, especially in cats with wet FIP (Fig. 14), and may be associated with jaundice and bilirubinuria. Hyperbilirubinemia in FIP is not caused by liver disease (Tasker, 2018), but rather by vasculitis, microhemorrhage, hemolysis, and destruction of damaged red blood cells by macrophages locally and in the liver. The released hemoglobin is finally metabolized to bilirubin, which is then conjugated in the hepatocytes and excreted in the urine. Glucuronidation is essential for bilirubin excretion, and genetic disorders affecting glucuronidation in humans prevent its excretion (Kalakonda et al., 2021). Cats as a species are deficient in the enzymes required for glucuronidation, making it difficult to excrete substances such as bilirubin (Court and Greenblatt 2000).
Although FIP can affect the kidneys and liver, it is not severe enough to cause significant loss of kidney or liver function. However, serum tests for blood urea nitrogen (BUN) and creatinine as indicators of kidney disease and alanine aminotransferase (ALT), alkaline phosphatase (ALP), and gamma glutamyltransferase (GGT) as indicators of liver disease are often mildly elevated in cats with FIP, especially with a more acute and serious disease (Fig. 14). Therefore, slightly abnormal test values should not be interpreted excessively if other clinical signs of liver or kidney disease are not present, while their significant increase should point to the possibility of concurrent and possibly predisposing diseases of these organs.
Serum can also be tested for other markers of systemic inflammation, such as increased levels of alpha-1-acid glycoprotein (AGP) (Paltrinieri et al., 2007) and feline serum amyloid A (fSAA) (Yuki et al., 2020). They may also prove useful in monitoring response to antiviral therapy (Krentz et al., 2021).
Radiography can be helpful in identifying chest and abdominal effusions. Abdominal ultrasound can reveal a smaller amount of effusion, identify enlarged mesenteric and ileo-cecal-colic lymph nodes, thickening of the colonic wall and lesions in organs such as the kidneys, liver and spleen (Lewis and O'Brien 2010). It may also be useful in examining the chest for lesions and assisting with needle aspiration or biopsy.
Antibody titers against FCoV have decreased since the first report nearly 50 years ago (Pedersen 1976b). The reference antibody test uses indirect fluorescent antibody staining (IFA) IFA titers ≥ 1:3200 in FIP cats are higher than most FECV-exposed cats (1:25–1:400). Newer tests often use ELISA procedures for rapid in-house or laboratory testing, but are qualitative rather than quantitative. IFA antibody titers decrease during successful antiviral treatment in many cats, but remain high in others (Dickinson et al., 2020; Krentz et al., 2021). Sequential titers can show a gradual increase in titers during the development of FIP (Pedersen et al., 1977), but previous serum samples are rarely available for comparison. Like most tests, FCoV antibody levels should not be used as the sole criterion to diagnose or rule out FIP (Felten and Hartmann, 2019) or to assess treatment success (Krentz et al., 2021).
Reverse transcriptase polymerase chain reaction (RT-PCR) is the primary means of identifying FCoV RNA in inflammatory effusions, fluids, or affected tissues (Felten and Hartmann, 2019). Accessory gene 7b RNA is present at the highest levels in tissues, fluids or exudates infected with FECV or FIPV, making it the most sensitive target for detecting low levels of virus (Gut et al., 1999). RT-PCR for FIPV S gene mutations is often used in samples that are positive for 7b RNA to be specific for FIPV (Felten et al., 2017). Other studies suggest that RT-PCR assays for FIPV-specific S gene mutations have similar specificity for FIP, but at the cost of a significant loss of sensitivity (Barker et al., 2017). A decrease in sensitivity is associated with an increase in the number of false negative results. False-negative RT-PCR tests also occur in samples that do not contain sufficient numbers of infected macrophages or in cats with very low levels of virus. False-negative results are especially common when testing whole blood.
Immunohistochemistry (IHC) detects feline coronavirus nucleocapsid protein in formalin-fixed tissues with high sensitivity and specificity, but is not as popular as RT-PCR (Litster et al., 2013; Ziółkowska et al., 2019). Specimens for IHC must contain intact infected macrophages (Fig. 17), which requires careful separation of cells from effusions and mounting them on slides, or formalin-fixed, paraffin-embedded diseased tissues that show lesions compatible with FIP. The coronavirus antigen in macrophages within a typical FIP lesion or fluid is seen only in FIP, giving IHC a high level of specificity.
Figure 17. Histological section from the thickened colon of a cat with the intestinal form of FIP. The thickened wall contained foci of macrophages (square area), which stained positive (brown-red) for FIPV nucleocapsid protein by immunoperoxidase.
A thorough ophthalmological examination is necessary to diagnose the characteristic changes of FIP (Pfeiffer and Wilcock 1991; Andrew, 2000). A sample of aqueous humor from the anterior chamber of an inflamed eye may also be useful for cytology, PCR and IHC.
Neurological FIP is often diagnosed using contrast-enhanced magnetic resonance imaging (MRI) and is often associated with cerebrospinal fluid (CSF) analysis (Crawford et al., 2017; Tasker, 2018; Dickinson et al., 2020). However, these are expensive procedures that are not always available and carry a certain risk for the cat. MRI lesions include obstructive hydrocephalus, syringomyelia, and herniation of the foramen magnum with contrast enhancement of the meninges of the brain and spinal cord and ependyma of the third ventricle, mesencephalic aqueduct, and brainstem. CSF shows an increased number of proteins and cells (neutrophils, lymphocytes, monocytes/macrophages) and, if present, can be reliable material for PCR or IHC examination.
Neurologic and/or ocular forms of FIP are often confused with systemic feline toxoplasmosis, and many cats with FIP are empirically treated for toxoplasmosis before a diagnosis of FIP is made. Fortunately, the availability of effective treatment for FIP has curtailed this practice. Systemic toxoplasmosis is much less prevalent than FIP, and fewer than 1 % cats with FIP were serologically positive in one field study.1 Therefore, testing or treatment for toxoplasmosis should only be considered once FIP has been adequately diagnosed.
Antiviral treatment as a diagnostic tool
Figure 18. A cat with FIP at the start of treatment with GS-441524 (a) and after 1 week (b). The answer is quick, the fever will disappear within 24-48 hours and the general state of health will improve significantly within 1-2 weeks. This type of response is often used to confirm a diagnosis of FIP.
Situations commonly occur where clinical findings point to FIP but doubts remain. Then there is a choice of performing several diagnostic tests, which may not lead to a more definitive diagnosis. An alternative diagnostic approach is treatment with a suitable antiviral for 1-2 weeks in the correct dose for the suspected form of FIP.2 Treatment often produces clinical improvement in as little as 24-48 hours and this rapidly progresses over the next 2 weeks and the total duration of treatment (Fig. 18). No response to test treatment and/or deterioration in health would indicate the need for further investigation of the cause(s) of ill health.
FIP Treatment
Before 2017, there was no cure for FIP, and treatment was mainly aimed at alleviating the symptoms of the disease (Izes et al., 2020). Such supportive treatment was aimed at maintaining good nutrition, controlling inflammation (corticosteroids), changing immune responses (interferons, cyclophosphamide, chlorambucil) and inhibiting key cytokine responses (pentoxifylline and other TNF-alpha inhibitors). Nutritional supplements that were supposed to help specific organ functions were also commonly used, such as one (Polyprenyl Immunostimulant) that was supposed to improve immunity and prolong survival in cats with dry but not wet FIP (Legendre et al., 2017). The effect of good supportive care on survival could not be determined because most cats were euthanized after diagnosis or within days or weeks. The survival rate for even the mildest forms of dry FIP and the most permanent treatment in one study was only 13 % at 200 days and 6 % at 300 days (Legendre et al., 2017).
Many commercially available drugs and compounds inhibit FIPV infection or replication in vitro, some of which are drugs known to inhibit specific HIV or hepatitis C proteins, while others work by inhibiting normal cellular processes that the virus usurps for its own life cycle (Hsieh et al., 2010; Izes et al., 2020; Delaplace et al., 2021). These various drugs and agents include cyclosporine and related immunophilins, several nucleoside and protease inhibitors, vioporin inhibitors, pyridine N-oxide derivatives, chloroquine and related compounds, ivermectin, several plant lectins, ubiquitin inhibitors, itraconazole, and several antibiotics. However, the concentrations required to inhibit viral replication in vitro often approach toxic values for cells. It has also been difficult to transfer favorable in vitro conclusions to animals, and studies in sick cats have rarely followed. Ribavarin inhibits FIPV replication in vitro, but was not effective as a treatment for experimental FIP (Weiss et al., 1993). The efficacy of chloroquine was tested in laboratory cats infected with FIPV, but clinical outcomes in treated cats were only slightly better than untreated ones and hepatotoxicity was demonstrated (Takano et al., 2013). A 3-month-old kitten with chest wet FIP treated with itraconazole and prednisolone developed neurological FIP and was euthanized after 38 days of treatment (Kameshima et al., 2020). Mefloquine also inhibited FIPV replication at low concentrations in cultured feline cells without cytotoxic effects, and preliminary pharmacokinetic studies in cats appeared favorable (Yu et al., 2020), but evidence of its safety and efficacy in clinical trials in cats with FIP has yet to be established. published.
A breakthrough in the treatment of FIP occurred in 2016-2019 when antiviral drugs were reported that target specific FIPV proteins essential for replication. The first of these drugs was GC376, a major protease inhibitor (Mpro ) FIPV (Kim et al., 2016; Pedersen et al., 2018). Protease inhibitors prevent the formation of individual viral proteins by inhibiting their cleavage from polyprotein precursors. GC376 was able to cure all experimentally infected cats and 7 of 21 cats with naturally occurring wet and dry FIP, but was less effective for cats with ocular or neurological signs (Pedersen et al., 2018). The second of these drugs was GS-441514, the active part of the prodrug remdesivir (Gilead Sciences; Murphy et al., 2018; Pedersen et al., 2019). GS-441524 is an adenosine nucleoside analog that blocks FIPV replication by inserting a nonsense adenosine into the developing viral RNA. GS-441524 was also able to cure all experimentally infected cats (Murphy et al., 2018) and 25/31 cats with naturally occurring wet and dry FIP (Pedersen et al., 2019). It has also been shown to be effective at higher doses in several cats with ocular and neurological FIP (Pedersen et al., 2019) and is now the drug of first choice for cats with neurological FIP (Dickinson et al., 2020). GS-441524 has cured thousands of FIP cats from around the world over the past three years, with an overall cure rate of just over 90 % (Jones et al., 2021).1
Although the ability of GC376 and GS-441524 to treat cats has been known for several years, neither is currently legally available in most countries. The rights to GC376 have been purchased by Anivive, but it has not yet been launched.3 Potential conflicts with the development of remdesivir for the treatment of COVID-19 in humans led Gilead Sciences to withhold rights to GS-441524 for animal use, prompting the creation of an unapproved source for GS-441524 from China (Jones et al, 2021).1,2,4 Remdesivir is rapidly metabolized in the body to GS-441524 and has been approved for the treatment of FIP in some countries.2 GS-441524 can also be administered orally in higher doses and is currently commonly used in practice (Krentz et al., 2021).1
The efficacy of drugs such as GC376 and GS-441524 on FIP cats, the use of which preceded the COVID-19 pandemic, has been recognized by researchers investigating related SARS-CoV 2 inhibitors (Yan et al., 2020; Vuong et al., 2021). Remdesivir, an injectable drug called glaucoma (Gilead), has been used worldwide to reduce mortality from COVID-19 (Beigel et al., 2020). GC373, the active form of the prodrug GC376, has undergone simple modifications to increase efficacy and oral bioavailability (Vuong et al., 2021). The GC373-related drug, nirmatrelvir, has been successfully tested against early COVID-19 infections and has been approved for the treatment of early COVID-19 and marketed as paxlovid (Pfizer). Paxlovid consists of two medicines, nirmatrevir and the HIV protease inhibitor ritonavir. Ritonavir is not a significant inhibitor of SARS-CoV 2, but is reported to prolong the half-life of Mprowhen used in combination (Vuong et al., 2020). Nirmatrelvir and paxlovid have not been tested in cats with FIP at present, but based on experience with the closely related drug GC376, oral treatment of some forms of FIP may be important in the future.
Two other nucleoside analogs, EIDD-1931 and EIDD-2801 (Painter et al., 2021), have been investigated for the treatment of multiple RNA virus infections in humans and animals. EIDD-1931 is the experimental designation for beta-D-N4-hydroxycytidine, a compound widely studied since the 1970's. Beta-D-N4-hydroxycytidine is metabolized to a ribonucleoside analog, which is incorporated into RNA instead of cytidine and leads to fatal mutations in the viral RNA strand. The compound is an inhibitor of a wide variety of human and animal RNA viruses, including all known coronaviruses. EIDD-1931 was modified to increase oral absorption and was termed EIDD-2801 (molnupiravir) (Painter et al., 2021). Molnupiravir is deesterified in the body to its active ingredient, beta-D-N4-hydroxycytidine. Therefore, EIDD-1931 and molnupiravir are analogous to GS-441524 and remdesivir. Molnupiravir is marketed for the home treatment of primary COVID-19 under the names Lagevrio (Merck, USA) or Molnulup (Lupine, India).
Both EIDD-1931 and EIDD-2801 have been shown to be effective in inhibiting FIPV in tissue culture (Cook et al., 2021), and EIDD-2801 is currently used to treat some cases of FIP in the field.5,7 The effective concentration of 50 % (EC50) for EIDD-1931 against FIPV is 0.09 µM, EIDD-2801 0.4 µM and GS-441524 0.66 µM (Cook et al., 2021). The percentage cytotoxicity at 100 μM for these compounds is 2.8, 3.8 and 0.0. Thus, EIDD-1931 and -2801 are slightly more inhibitory to viruses, but more cytotoxic than GS-441524. Resistance to GS-441524 has been reported in some cases of FIP (Pedersen et al., 2019) and to remdesivir in patients with COVID-19 (Painter et al., 2021), but these isolates remain sensitive to molnupiravir (Sheahan et al., 2020). This may prove useful in combating resistance to GS-441524 in cats and humans and in developing multidrug therapy to prevent the development of resistance.
What will full approval of medicines like molnupiravir and paxlovid mean for cats? Full human approval should allow veterinarians in most countries to legally procure medicinal products authorized for human consumption for direct use in animals, provided that the guidelines for use in non-food producing animals are followed.6 This requires a reformulation of a medicine made for humans and purchased at a price for humans. Hopefully, antivirals similar or identical to those approved for humans will be licensed exclusively for animals and sold at a much lower price, but this is likely to take years.
Commercial and policy issues that limit the current use of antivirals such as GS-441524 in animal diseases such as FIP are for current cat owners and feline support groups who have already bypassed the current drug approval system and its emphasis on overriding human needs, irrelevant (Jones et al., 2021; Krentz et al., 2021). Advocates of FIP treatment are currently found around the world and often associate under the expanded FIP Warrior brand. Members of these groups often act as intermediaries between owners, veterinarians and antiviral suppliers and often provide advice to those who are unable to obtain veterinary treatment assistance. Some of these groups, such as FIP Warriors Czech Republic / Slovakia7, have placed their experience with FIP treatment on the Internet, where they provide much-needed information about current antiviral treatment.
Current situation of FIP treatment
The current drug of choice for the treatment of FIP is the adenosine nucleoside analog GS-441524, which was first published in the scientific literature under experimental conditions (Murphy et al., 2018) and later against naturally occurring disease (Pedersen et al., 2019). Although initial experimental and field studies of GS-441524 were conducted in collaboration between researchers at Gilead Sciences and the University of California, Davis, the relationship between Remdesivir and GS-441524 and the onset of the COVID-19 pandemic in 2019 led Gilead Sciences to eventually did not grant rights to use GS-441524 to animals on the grounds that it could interfere with the development of Remdesivir for human use.4 Objections to this decision have been raised directly by the company and in several internet forums.4 Subsequent pressure from cat owners, cat rescue groups and cat lovers, along with opportunistic Chinese drug manufacturers, quickly created an alternative unapproved source of GS-441524, its market and treatment network.4 This network has largely bypassed veterinarians, most of whom have decided to wait for the drug to be legalized (Jones et al., 2021). The result of this relationship was an almost seamless transition of FIP treatment with GS-441524 from the laboratory to a rapidly expanding worldwide network of groups, under the umbrella of FIP Warriors (Jones et al., 2021).4,7
The sale and use of GS-441524 in practice for the treatment of FIP began almost immediately with the first publication of the results of field trials (Pedersen et al., 2019) (Fig. 19).
Figure 19. Graph of the monthly development of treatment of cats from the Czech Republic and Slovakia since August 2019. This graph comes from the FIP Warrior CZ / SK website.1 These figures reflect the experiences of other FIP Warrior groups around the world. Since 2019, when the first field study GS-441524 was published (Pedersen et al. 2019), thousands of cats have been successfully treated for FIP worldwide. The winter peaks of the disease reflect a late spring and summer increase in the number of kittens born and a high incidence of FIP, which usually begins at 3 to 6 months of age (Fig. 6). This chart is from the FIP Warrior CZ / SK website.1
Figure 20. The main participants in the GS-441524 treatment. This chart is from the FIP Warriors CZ / SK website.1
The fact that GS-441524 is not legally approved for use in animals has prevented many veterinarians from recognizing or participating in this treatment. Only 25 % cats in the CZ / SK treated group received veterinary support during treatment (Fig. 20), although more veterinarians may have been involved in the diagnosis of the disease. Interestingly, this number was higher than the 8.7 % treated cats in the United States that received veterinary care (Jones et al., 2021). However, participants in CZ / SK studies and similar groups around the world are not without medical experience, as many of them are engaged in temporary care / rescue and have had considerable direct and indirect veterinary experience with cat diseases and their treatment and castration programs.
From the first laboratory studies and research of Chinese manufacturers, it was known that GS-441524 can be absorbed orally, although with less efficiency (Kim et al. 2016).9 The first sellers of GS-441524 further investigated this fact and found that effective blood levels could be achieved by increasing the amount administered orally compared to injection.8 Supplements have often been added to GS-441524 oral capsules or tablets, claiming that they increase absorption or have additive therapeutic benefits (Krentz et al., 2011). Most major retailers of GS-441524 now offer oral versions, and oral therapy is becoming increasingly popular either as a single treatment or in combination with GS-441524 (Figure 21). The success of GS-441524 oral therapy did not differ significantly from GS-441524 injection therapy (Figure 22).
Figure 21. Comparison of the use of oral (tablets or capsules) and injectable (subcutaneous) forms of GS-441524 for the treatment of FIP in cats from the Czech Republic and Slovakia. This chart is from the FIP Warriors CZ / SK website.1
Figure 22. There is no significant difference in treatment success with oral administration of GS-441524 compared to subcutaneously administered GS, but the actual amount (mg) of drug administered orally in each dose is up to twice the amount contained in the same dose of GS injection. This chart is from the FIP Warriors CZ / SK.1 website
The recommended dosing schedule for GS-441524 based on published data from field studies (Pedersen et al., 2019) was 4 mg / kg, subcutaneous (SC), daily (q24h), ie 4 mg / kg, SC, q24h. This recommended starting dose for cats with wet or dry FIP without ocular or neurological symptoms tended to increase to 6 mg / kg SC q24h over time (Fig. 23). 8 mg / kg SC q24h is the current recommended dose for cats with ocular symptoms and 10 or 12 mg / kg SC q24h for cats with neurological symptoms.
Figure 23. Daily dose of GS-441524, which was used to treat FIP in cats from the Czech Republic and Slovakia. The usual starting dose was 6 mg / day, with some cats requiring higher doses based on response to treatment, form of the disease and recurrence after treatment appeared to be successful. GS-441524 oral formulations are usually labeled to match the dosage used for the injectable drug, but contain up to twice the labeled amount. This chart is from the FIP Warrior CZ / SK website.1
The optimal duration of treatment, as determined in the initial clinical study, is 84 days (Pedersen et al., 2019). In some cases of acute wet FIP in younger cats, healing has been achieved in 6-8 weeks, but some cats need more than 84 days. As shown in Figure 24.72 % cats were treated for 81-90 days, 19 % longer and only 9 % were treated shorter. Unfortunately, there is no simple and accurate test to determine the moment of cure, and the decision to stop treatment is based on a complete return to health and normal blood test values. Cats treated for much longer than 100 days were usually those requiring a GS dose higher than 12 mg / kg per day by injection or equivalent oral dose, cats that relapsed during the 12-week post-treatment observation period, cats with neurological disease or cats that have become resistant to GS-441524.
Figure 24. Duration of treatment with GS-141524 in 352 cats successfully treated for all forms of FIP. This chart is from the FIP Warriors CZ / SK website.1
Figure 25. Initial treatment was successful in 88.1 % cats and 6.2 % cats died or were sacrificed either due to insufficient response to treatment, financial reasons or treatment side effects. Another 5.7 % cats relapsed after initial treatment and approximately the same number of cats either recovered or died after further treatment. This chart is from the FIP Warriors CZ / SK.1 website
The treatment success rate for all forms of FIP in cats from the Czech Republic and Slovakia is 88.1 % in the first treatment, but when cats that relapsed after the first treatment and recovered after the second treatment (3.1 %) were included, the overall success rate was more as 91 % (Fig. 25). This cure rate is identical to the cure rate of other groups of FIP fighters (Jones et al., 2021). Treatment success did not differ between cats with wet or dry FIP and without ocular or neurological impairment (Fig. 26). However, the cure rate in cats with ocular and neurological impairments was lower, at 80 % compared to 92 % in all other forms of FIP (Fig. 26).
Figure 26. Cure rate of cats with wet or dry FIP without ocular or neurological symptoms and cats with ocular or neurological disease as the main feature of their disease. This chart is from the FIP Warriors CZ / SK website.1
Figure 27. The condition of the cats one year after the successful completion of treatment with GS-441524. This chart is from the FIP Warriors CZ / SK website.1
Cats that have been successfully treated for FIP have been followed for 4 to 5 years, including cases reported in the first field studies. There have been no recurrences or recurrent cases of FIP in this group of first field trials. Data on annual survival are available from a much larger population of the CZ / SK study, which shows that 90.5 % cats are still healthy one year after the end of treatment (Fig. 27). Only 1.3 % of these cats died from causes other than FIP and 8.2 % cohort is currently in an unknown medical condition. The low proportion of cats that died of unknown causes within a year of treatment and their positive response to treatment suggest that FIP has been diagnosed correctly.
EIDD-2801 (molnupiravir) is currently being used in the field for the main treatment and for the treatment of GS-441524-resistant cats.5,7,9 EIDD-1931, the active form of EIDD-2081, needs to be further researched because it is no longer covered by patent protection and is thus easily approved for use in animals if it is found to be truly safe and effective.5 Nirmatrelvir, an oral form of GC373 and a closely related GC376, still needs to be studied for the treatment of FIP.
Acknowledgement
I am indebted to Ladislav Mihok and his collaborators at “FIP Warriors Czech Republic/Slovakia” for allowing me to share data from their website. This website contains the most significant, thorough and organized collection of data on the treatment of FIP with antivirals to date. The website also contains useful information and advice on initiating, managing and monitoring current treatment. The collection of cats and their data is continuously and regularly updated and at the time of writing included over 600 cats with FIP.
References
Addie DD, Toth S, Murray GD, Jarrett O, 1995. Risk of feline infectious peritonitis in cats naturally infected with feline coronavirus. American Journal of Veterinary Research, 56, 429-34.
Addie DD, Schaap IA, Nicolson L, Jarrett O, 2003. Persistence and transmission of natural type I feline coronavirus infection. Journal of General Virology 84, 2735–2744.
Andrew SE, 2000. Feline infectious peritonitis. Veterinary Clinics of North America and Small Animal Practice 30, 987-1000.
Barker EN, Stranieri A, Helps CR, Porter EL, Davison AD, Day MJ, Knowles T, Kipar A, Tasker S, 2017. Limitations of using feline coronavirus spike protein gene mutations to diagnose feline infectious peritonitis. Veterinary Research 48, 60.
Beigel JH, Tomashek KM, Dodd LE, Mehta EK, Zingman BS, et al., 2020. Remdesivir for the Treatment of Covid-19 - Final Report. New England Journal of Medicine, 383, 1813-1826,
Bubenikova J, Vrabelova J, Stejskalova K, Futas J, Plasil M, Cerna P, Oppelt J, Lobova D, Molinkova D, Horin P, 2020. Candidate gene markers associated with fecal shedding of the feline enteric coronavirus (FECV). Pathogens 9, 958.
Cassado Ados A, D'Império Lima, Bortoluci KR., 2015. Revisiting mouse peritoneal macrophages: heterogeneity, development, and function. Frontiers in Immunology 6, 225.
Cave TA, Thompson H, Reid SW, Hodgson DR, Addie DD, 2002. Kitten mortality in the United Kingdom: a retrospective analysis of 274 histopathological examinations (1986 to 2000). Veterinary Record 151, 497–501.
Cook SE, Vogel H, Castillo D, Olsen M, Pedersen N, Murphy BG, 2021. Investigation of monotherapy and combined anticoronaviral therapies against feline coronavirus serotype II in vitro. Journal of Feline Medicine and Surgery. doi: 10.1177 / 1098612X211048647. Epub ahead of print. PMID: 34676775.
Cornelissen E, Dewerchin HL, Van Hamme E, Nauwynck HJ, 2007. Absence of surface expression of feline infectious peritonitis virus (FIPV) antigens on infected cells isolated from cats with FIP. Veterinary Microbiology. 121, 131-137,
Cotter SM, Gilmore CE, Rollins C. 1973, Multiple cases of feline leukemia and feline infectious peritonitis in a household. Journal of the American Veterinary Medical Association 162, 1054–1058.
Court MH., Greenblatt DJ. 2000, Molecular genetic basis for deficient acetaminophen glucuronidation by cats: UGT1A6 is a pseudogene, and evidence for reduced diversity of expressed hepatic UGT1A isoforms Pharmacogenetics, 10, 355-369
Crawford AH, Stoll AL, Sanchez-Masian D, Shea A, Michaels J, Fraser AR, Beltran E, 2017. clinicopathologic features and magnetic resonance imaging findings in 24 cats with histopathologically confirmed neurologic feline infectious peritonitis. Journal of Veterinary Internal Medicine 31, 1477–1486.
Day MJ, 2010. Aging, immunosenescence and inflammageing in the dog and cat. Journal of Comparative Pathology 142 Suppl 1, S60-69.
Delaplace M, Huet H, Gambino A, Le Poder S, 2021. Feline coronavirus antivirals: A review. Pathogens 10, 1150. doi: 10.3390 / pathogens10091150.
Dempsey SM, Ewing PJ, 2011. A Review of the Pathophysiology, Classification, and Analysis of Canine and Feline Cavitary Effusions. Journal of the American Animal Hospital Association 47, 1–11.
de Sousa JR, Sotto MN, Simões Quaresma JA, 2017. Leprosy as a complex infection: Breakdown of the Th1 and Th2 immune paradigm in the immunopathogenesis of the disease. Frontiers in Immunology 8,1635.
Dewerchin HL, Cornelissen E, Van Hamme E, Smits K, Verhasselt B, Nauwynck HJ, 2008. Surface-expressed viral proteins in feline infectious peritonitis virus-infected monocytes are internalized through a clathrin- and caveolae-independent pathway. Journal of General Virology 89, 2731–2740
Dewerchin HL, Desmarets LM, Noppe Y, Nauwynck HJ, 2014. Myosins 1 and 6, myosin light chain kinase, actin and microtubules cooperate during antibody-mediated internalization and trafficking of membrane-expressed viral antigens in feline infectious peritonitis virus infected monocytes. Veterinary Research 45, 17.
Dickinson PJ, Bannasch M, Thomasy SM, Murthy VD, Vernau KM, Liepnieks M, Montgomery E, Knickelbein KE, Murphy B, Pedersen NC, 2020. Antiviral treatment using the adenosine nucleoside analogue GS ‐ 441524 in cats with clinically diagnosed neurological feline infectious peritonitis. Journal of Veterinary Internal Medicine 34, 1587–1593.
Drechsler Y, Alcaraz A, Bossong FJ, Collisson EW, Diniz PP, 2011. Feline coronavirus in multicat environments. Veterinary Clinics North America and Small Animal Practice41, 1133-1169.
Fankauser R, Fatzer R, 1997. Meningitis and chorioependymitis granulomatosa der Katze. Possible conditions for infectious peritonitis (FIP). Client Practice 22, 19–22.
Felten S, Leutenegger CM, Balzer HJ, Pantchev N, Matiasek K, Wess G, Egberink H, Hartmann K, 2017. Sensitivity and specificity of a real-time reverse transcriptase polymerase chain reaction detecting feline coronavirus mutations in effusion and serum / plasma of cats to diagnose feline infectious peritonitis. BMC Veterinary Research 13, 228.
Felten S, Hartmann K, 2019. Diagnosis of Feline Infectious Peritonitis: A Review of the Current Literature. Viruses 11, 1068.
Fischer Y, Sauter-Louis C, Hartmann K, 2012. Diagnostic accuracy of the Rivalta test for feline infectious peritonitis. Veterinary Clinical Pathology 41, 558–67.
Foley JE, Poland A, Carlson J, Pedersen NC, 1997. Risk factors for feline infectious peritonitis among cats in multiple-cat environments with endemic feline enteric coronavirus. Journal of the American Veterinary Medicine Association 210, 1313-1318.
Foley JE, Lapointe JM, Koblik P, Poland A, Pedersen NC, 1998. Diagnostic features of clinical neurologic feline infectious peritonitis. Journal of Veterinary Internal 12, 415–423.
Gaskell RM, Povey RC, 1977. Experimental induction of feline viral rhinotracheitis virus re-excretion in FVR-recovered cats. Veterinary Record 100, 128–133.
Golovko L, Lyons LA, Liu H, Sørensen A, Wehnert S, Pedersen NC, 2013. Genetic susceptibility to feline infectious peritonitis in Birman cats. Virus Research 175, 58–63.
Gunn-Moore DA, Gaunt C, Shaw DJ, 2012. Incidence of mycobacterial infections in cats in great britain: estimate from feline tissue samples submitted to diagnostic laboratories. Transboundary and Emerging Diseases. 60, 338-344.
Gut, M, Leutenegger, CM, Huder, JB, Pedersen NC, H, 1999. One-tube fluorogenic reverse transcription-polymerase chain reaction for the quantitation of feline coronaviruses. Journal of Virological Methods 77, 37–46.
Hardy WD Jr., 1981. Feline leukemia virus non-neoplastic diseases. Journal of the American Animal Hospital Association 17, 941-949.
Healey EA, Andre NM, Miller AD, Whitaker GR, Berliner EA, 2022. Outbreak of feline infectious peritonitis (FIP) in shelter-housed cats: Molecular analysis of the feline coronavirus S1/S2 cleavage site consistent with a 'circulating virulent-avirulent theory' of FIP pathogenesis. Journal of Feline Medicine and Surgery Open Reports 8, 20551169221074226.
Herrewegh AAPM, Mähler M, Hedrich HJ, Haagmans BL, Egberink HF, Horzinek MC, Rottier PJM, de Groot RJ, 1997. Persistence and evolution of feline coronavirus in a closed cat-breeding colony. Virology 234, 349–363.
Herrewegh AA, Smeenk I, Horzinek MC, Rottier PJ, de Groot RJ, 1998. Feline coronavirus type II strains 79-1683 and 79-1146 originate from a double recombination between feline coronavirus type I and canine coronavirus. Journal of Virology 72, 4508–4514.
Hickman MA, Morris JG, Rogers QR, Pedersen NC, 1995. Elimination of feline coronavirus infection from a large experimental specific pathogen-free cat breeding colony by serologic testing and isolation, Feline Practice 23, 96–102.
Hsieh LE, Lin CN, Su BL, Jan TR, Chen CM, Wang CH, Lin DS, Lin CT, Chueh LL. 2010. Synergistic antiviral effect of Galanthus nivalis agglutinin and nelfinavir against feline coronavirus. Antiviral Research 88, 25–30.
Holzworth J, 1963. Some important disorders of cats. Cornell Veterinarian 53, 157–160.
Izes AM, Yu J, Norris JM, Govendir M, 2020. Current status on treatment options for feline infectious peritonitis and SARS-CoV-2 positive cats. Veterinary Quarterly 40, 322–330.
Jones S, Novicoff W, Nadeau J, Evans S, 2021. Unlicensed GS-441524-like antiviral therapy can be effective for at-home treatment of feline infectious peritonitis. Animals 11, 2257.
Kalakonda A, Jenkins BA, John S. Physiology, Bilirubin. [Updated 2021 Sep 16]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2022 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK470290/
Mustaffa-Kamal F, Liu H, Pedersen NC, Sparger EE, 2019. Characterization of antiviral T cell responses during primary and secondary challenge of laboratory cats with feline infectious peritonitis virus (FIPV). BMC Veterinary Research 15,165.
Kameshima S, Kimura Y, Doki T, Takano T, Park CH, Itoh N, 2020. Clinical efficacy of combination therapy of itraconazole and prednisolone for treating effusive feline infectious peritonitis. Journal of Veterinary Medical Science 82, 1492–1496.
Kim Y, Liu H, Galasiti Kankanamalage AC, Weerasekara S, Hua DH, Groutas WC, Chang KO, Pedersen NC, 2016. Reversal of the progression of fatal coronavirus Infection in cats by a broad-spectrum coronavirus protease inhibitor. PLoS Pathogens 12: e1005531.
Kipar A, May H, Menger S, Weber M, Leukert W, Reinacher M, 2005. Morphologic features, and development of granulomatous vasculitis in feline infectious peritonitis. Veterinary Pathology 42, 321–330.
Krentz D., Zenger K., Alberer M., Felten S., Bergmann M, Dorsch R., Matiasek, K., Kolberg, L., Hofmann-Lehmann, R., Meli, ML, et al., 2021. Curing cats with feline infectious peritonitis with an oral multi-component drug containing GS-441524. Viruses 13, 2228.
Legendre AM, Kuritz T, Galyon G, Baylor VM, Heidel RE, 2017. Polyprenyl immunostimulant treatment of cats with presumptive non-effusive feline infectious peritonitis in a field study. Frontiers in Veterinary Science 4, 7.
Teacher C, Lê-Bury P, Pizarro-Cerdá J, Dussurget O, 2020. Emerging Evasion Mechanisms of Macrophage Defenses by Pathogenic Bacteria. Frontiers in Cellular and Infection Microbiology, 10, 538.
Lewis KM, O'Brien RT, 2010. Abdominal ultrasonographic findings associated with feline infectious peritonitis: a retrospective review of 16 cases. Journal of the American Animal Hospital Association. 46, 152-60.
Licitra BN, Millet JK, Regan AD, Hamilton BS, Rinaldi VD, Duhamel GE, Whittaker GR, 2013. Mutation in spike protein cleavage site and pathogenesis of feline coronavirus. Emerging Infectious Diseases 19, 1066–1073.
Lin CN, Su BL, Wang CH, Hsieh MW, Chueh TJ, Chueh LL, 2009. Genetic diversity and correlation with feline infectious peritonitis of feline coronavirus type I and II: A 5-year study in Taiwan. Veterinary Microbiology 136, 233–239.
Litster AL. Pogranichniy R, Lin TL, 2013. Diagnostic utility of a direct immunofluorescence test to detect feline coronavirus antigen in macrophages in effusive feline infectious peritonitis. Veterinary Journal 198, 362–366.
Lloret A, Hartmann K, Pennisi MG, Ferrer L, Addie D, Belák S, Boucraut-Baralon C, Egberink H, Frymus T, Gruffydd-Jones T, et al., 2013. Rare systemic mycoses in cats: blastomycosis, histoplasmosis and coccidioidomycosis: ABCD guidelines on prevention and management. Journal of Feline Medicine and Surgery 15, 624–627.
Longstaff L, Porter E, Crossley VJ, Hayhow SE, Helps CR, Tasker S, 2017. Feline coronavirus quantitative reverse transcriptase polymerase chain reaction on effusion samples in cats with and without feline infectious peritonitis. Journal of Feline Medicine and Surgery 19, 240–245.
Mahase E. 2021. Covid-19: Molnupiravir reduces risk of hospital admission or death by 50% in patients at risk, MSD reports. BMJ 375, n2422.
Malbon AJ, Meli ML, Barker EN, Davidson AD, Tasker S, Kipar A, 2019. inflammatory mediators in the mesenteric lymph nodes, site of a possible intermediate phase in the immune response to feline coronavirus and the pathogenesis of feline infectious peritonitis? Journal of Comparative Pathology 166, 69–86.
Malbon AJ, Russo G, Burgener C, Barker EN, Meli ML, Tasker S, Kipar A, 2020. the effect of natural feline coronavirus infection on the host immune response: A whole-transcriptome analysis of the mesenteric lymph nodes in cats with and without feline infectious peritonitis. Pathogens 7, 524.
Mor G, Cardenas I, 2010. The immune system in pregnancy: A unique complexity. American Journal of Reproductive Immunology 63, 425–433.
Murphy BG, Perron M, Murakami E, Bauer K, Park Y, Eckstrand C, Liepnieks M, Pedersen NC, 2018. The nucleoside analog GS-441524 strongly inhibits feline infectious peritonitis (FIP) virus in tissue culture and experimental cat infection studies. Veterinary Microbiology 219, 226–233.
Painter WP, Holman W, Bush JA, Almazedi F, Malik H, Eraut NCJE, Morin MJ, Szewczyk LJ, Painter GR, 2021. Human safety, tolerability, and pharmacokinetics of molnupiravir, a novel broad-spectrum oral antiviral agent with activity against SARS-CoV-2. Antimicrobial Agents and Chemotherapeutics 65: e02428-20.
Paltrinieri S, Giordano A, Tranquillo V, Guazzetti S, 2007. Critical assessment of the diagnostic value of feline α1-acid glycoprotein for feline infectious peritonitis using the likelihood ratios approach. Journal of Veterinary Diagnostic Investigation. 19, 266-272.
Pearson M, LaVoy A, Evans S, Vilander A, Webb C, Graham B, Musselman E, LeCureux J, VandeWoude S, Dean GA, 2019. Mucosal Immune Response to Feline Enteric Coronavirus Infection. Viruses 11, 906.
Pedersen NC, 1976b. Serologic Studies of Naturally Occurring Feline Infectious
peritonitis. American Journal of Veterinary Research 37, 1447–1453.
Pedersen NC, 2009. A review of feline infectious peritonitis virus infection: 1963-2008. Journal of Feline Medicine and Surgery 11, 225–258.
Pedersen NC, Boyle J, 1980. Immunologic Phenomena in the Effusive Form of Feline Infectious Peritonitis. American Journal of Veterinary Research 41: 868–876.
Pedersen NC, Ward J, Mengeling WL, 1978. Antigenic relationship of the feline infectious peritonitis virus to coronaviruses of other species. Archives of Virology58, 45–53.
Pedersen NC, Allen CE, Lyons LA, 2008. Pathogenesis of feline enteric coronavirus infection. Journal of Feline Medicine and Surgery 10, 529–541.
Pedersen NC, Theilen G, Keane MA, Fairbanks L, Mason T, Orser B, Che CH, Allison C, 1977. Studies of naturally transmitted feline leukemia virus infection. American Journal of Veterinary Research 38, 1523–1531.
Pedersen NC, Boyle JF, Floyd K, Fudge A, Barker J, 1981. An enteric coronavirus infection of cats and its relationship to feline infectious peritonitis. American Journal of Veterinary Research 42, 368-377.
Pedersen NC, Meric SM, Hoe E, Johnson L. Plucker S, Theilen GH, 1982. The clinical significance of latent feline leukemia virus infection. Feline Practice 14, 32–48.
Pedersen NC, Black JW, Boyle JF, Evermann JF, McKeirnan AJ, Ott RL, 1984. Pathogenic differences between various feline coronavirus isolates. Advances in Experimental Medicine and Biology 173, 365–380.
Pedersen NC, Liu H, Dodd KA, Pesavento PA, 2009. Significance of coronavirus mutants in feces and diseased tissues of cats suffering from feline infectious peritonitis. Viruses1, 166-184.
Pedersen NC, Liu H, Durden M, Lyons LA, 2016. Natural resistance to experimental feline infectious peritonitis virus infection is decreased rather than increased by positive genetic selection. Veterinary Immunology and Immunopathology 171, 17–20.
Pedersen NC, Liu H, Scarlett J, Leutenegger CM, Golovko L, Kennedy H, Kamal FM, 2012. Feline infectious peritonitis: role of the feline coronavirus 3c gene in intestinal tropism and pathogenicity based upon isolates from resident and adopted shelter cats. Virus Research 165,17-28
Pedersen NC, Kim Y, Liu H, Galasiti Kankanamalage AC, Eckstrand C, Groutas WC, Bannasch M, Meadows JM, Chang KO, 2018. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. Journal of Feline Medicine and Surgery 20, 378–392.
Pedersen NC, Perron M, Bannasch M, Montgomery E, Murakami E, Liepnieks M, Liu H, 2019. Efficacy and safety of the nucleoside analog GS-441524 for the treatment of cats with naturally occurring feline infectious peritonitis. Journal of Feline Medicine and Surgery 21, 271–281.
Peiffer RL Jr, Wilcock BP, 1991. Histopathologic study of uveitis in cats: 139 cases (1978-1988). Journal of the American Veterinary Medical Association 198, 135–138.
Pesteanu-Somogyi LD, Radzai C, Pressler BM, 2006. Prevalence of feline infectious peritonitis in specific cat breeds. Journal of Feline Medicine and Surgery 8, 1–5.
Poland AM, Vennema H, Foley JE, Pedersen NC, 1996. Two related strains of feline infectious peritonitis virus isolated from immunocompromised cats infected with the feline enteric coronavirus. Journal of Clinical Microbiology 34, 3180–3184.
Regan A, Whitaker G, 2008. Utilization of DC-SIGN for entry of feline coronaviruses into host cells. Journal of Virology 82, 11992-11996.
Riemer F, Kuehner KA, Ritz S, Sauter-Louis C, Hartmann K, 2016. Clinical and laboratory features of cats with feline infectious peritonitis – a retrospective study of 231 confirmed cases (2000-2010). Journal of Feline Medicine and Surgery 18, 348–356.
Rohrbach BW, Legendre AM, Baldwin CA, Lein DH, Reed WM, Wilson RB, 2001. Epidemiology of feline infectious peritonitis among cats examined at veterinary medical teaching hospitals. Journal of the American Veterinary Medical Association 218, 1111–1115.
Rojko J, Hoover E, Quackenbush, S. Olsen RG, 1982. Reactivation of latent feline leukemia virus infection. Nature 298, 385–388.
Sheahan TP, Sims AC, Zhou S, Graham RL, Pruijssers AJ, Aostini, ML, Leist, SR, Schäfer, A, Dinnon, KH 3rd., Stevens, LJ et al., 2020. An orally bioavailable broad-spectrum antiviral inhibitions SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Science Translational Medicine. 12, eabb5883.
Spencer, SE, Knowles, T, Ramsey, IK. 2017. Pyrexia in cats. retrospective analysis of signalment, clinical investigations, diagnosis and influence of prior treatment in 106 referred cases. Journal of Feline Medicine and Surgery 19, 1123–1130.
Stella J, Croney C, Buffington T, 2013. Effects of stressors on the behavior and physiology of domestic cats. Applied Animal Behavior Science 143, 157-163.
Stranieri A, Giordano A, Bo S, Braghiroli C, Paltrnieri S, 2017. Frequency of electrophoretic changes consistent with feline infectious peritonitis in two different time periods (2004–2009 vs 2013–2014). Journal of Feline Medicine and Surgery 19, 880–887.
Takano T, Katoh Y, Doki T, Hohdatsu T, 2013. Effect of chloroquine on feline infectious peritonitis virus infection in vitro and in vivo. Antiviral Research. 99, 100–107.
Tasker S, 2018. Diagnosis of feline infectious peritonitis: Update on evidence supporting available tests. Journal of Feline Medicine and Surgery 20, 228–243.
Tekes G, Ehmann R, Boulant S, Stanifer ML, 2020. Development of feline ileum- and colon-derived organoids and their potential use to support feline coronavirus infection. Cells 9, 2085.
Terada Y, Matsui N, Noguchi K, Kuwata R, Shimoda H, Soma T, Mochizuki M, Maeda K, 2014. Emergence of pathogenic coronaviruses in cats by homologous recombination between feline and canine coronaviruses. PLoS One 9, e106534.
Van Hamme E, Dewerchin HL, Cornelissen E, Verhasselt B, Nauwynck HJ, 2008. Clathrin- and caveolae-independent entry of feline infectious peritonitis virus in monocytes depends on dynamin. Journal of General Virology 89, 2147–2156.
Vennema H, Poland A, Foley J, Pedersen NC, 1995. Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology 243, 150–157.
Vogel L, Van der Lubben M,, Te Lintelo EG, Bekker CPJ, Geerts T, Schuif LS, Grinwis GCM, Egberink HF, Rottier PJM, 2010. Pathogenic characteristics of persistent feline enteric coronavirus infection in cats. Veterinary Research 41, 71.
Vuong, W, Fischer C, Khan MB, van Belkum MJ, Lamer T, Willoughby, KD, Lu, J, Arutyenova, E, Joyce, MA, Saffran, HA et al., 2021. Improved SARS-CoV-2 Mpro inhibitors based on feline antiviral drug GC376: Structural enhancements, increased solubility, and micellar studies. European Journal of Medicinal Chemistry, 222, 113584.
Wang YT, Su BL, Hsieh LE, Chueh LL, 2013. An outbreak of feline infectious peritonitis in a Taiwanese shelter: Epidemiologic and molecular evidence for horizontal transmission of a novel type II feline coronavirus. Veterinary Research, 44, 57.
Ward JM, 1970. Morphogenesis of a virus in cats with experimental feline infectious peritonitis. Virology 41, 191–194.
Watanabe R, Eckstrand C, Liu H, Pedersen NC, 2018. Characterization of peritoneal cells from cats with experimentally-induced feline infectious peritonitis (FIP) using RNA-seq. Veterinary Research 49, 81.
Weiss RC, Scott FW, 1981. Antibody-mediated enhancement of disease in feline infectious peritonitis: comparisons with dengue hemorrhagic fever. Comparative Immunology, Microbiology and Infectious Diseases 4, 175-189.
Weiss RC, Cox NR, Martinez ML, 1993. Evaluation of free or liposome-encapsulated ribavirin for antiviral therapy of experimentally induced feline infectious peritonitis. Research in Veterinary Science 55, 162e72.
Wolfe, LG, Griesemer, RA, 1971. Feline infectious peritonitis: a review of gross and histopathologic lesions. Journal of the American Veterinary Medical Association 158, 987–993.
Worthing KA, Wigney DI, Dhand NK, Fawcett A, McDonagh P, Malik R, Norris JM, 2012. Risk factors for feline infectious peritonitis in Australian cats. Journal of Feline Medicine and Surgery 14, 405–412.
Yan VC, Muller FL, 2020. Advantages of the Parent Nucleoside GS-441524 over Remdesivir for Covid-19 Treatment. ACS Medicinal Chemistry Letters 11, 1361–1366
Yu J, Kimble B, Norris JM, Govendir M, 2020. Pharmacokinetic profile of oral administration of mefloquine to clinically normal cats: A preliminary in-vivo study of a potential treatment for feline infectious peritonitis (FIP). Animals 10, 1000.
Yuki M, Aoyama R, Nakagawa M, Hirano T, Naitoh E, Kainuma D, 2020. A Clinical Investigation on serum amyloid A concentration in client-owned healthy and diseased cats in a primary care animal hospital. Veterinary Sciences, 7, 45.
Ziółkowska N, Paździor-Czapula K, Lewczuk B, Mikulska-Skupień E, Przybylska-Gornowicz B, Kwiecińska K, Ziółkowski H, 2017. Feline infectious peritonitis: immunohistochemical features of ocular inflammation and the distribution of viral antigens in the structures of the eye. Veterinary Pathology, 54, 933-940.
Zook BC, King NW, Robinson RL, McCombs HL, 1968. Ultrastructural evidence for the viral etiology of feline infectious peritonitis. Veterinary Pathology 5, 91–95.
Hughes D, Howard G, Malik R, 2021. Treatment of FIP in cats with Remdesivir. Clinical review, 2021. The Veterinarian. https://www.turramurravet.com.au/wp-content/uploads/2021/07/FIP-Article_The-Veterinarian.pdf (Accessed 5 March 2022).
Anonymous. Thanks to Cats, One Promising Coronavirus Treatment is Already in Development-The GC376 story. 2021, https://anivive.com/coronavirus (Accessed 4 April 2022)
Abstract: Feline infectious peritonitis, or simply FIP, is a coronavirus viral disease in cats, usually less than three years old. It is manifested by an extreme inflammatory reaction in the tissues in the abdomen, kidneys and brain. This review article discusses the various diagnostic tests and their benefits in diagnosing cases of suspected FIP for definitive diagnosis. This review can help compare different diagnostic parameters and also raise awareness of their advantages and disadvantages.
Feline infectious peritonitis (FIP) is a well-known and widespread systemic coronavirus (CoV) disease in cats, characterized by fibrinous-granulomatous serositis with protein-rich effusions into body cavities, granulomatous necrotizing phlebitis and periphlebitis, and granulomatous inflammatory lesions in multiple organs (Weiss and Scott 1981; Kipar et al. 2005). Feline CoV (FCoV) is transmitted by the fecal-oral route and primarily infects enterocytes (Pedersen 1995), but subsequently spreads systemically through monocytic viremia (Meli et al 2004; Kipar et al 2005). It has been shown that increased viral replication capacity may be a key feature in the development of FIP, and it is also thought that FIP is caused by mutations in the common feline enteric coronavirus (FECV), which is found in cats worldwide and is not a serious infection (Pedersen et al 2009; Healey et al 2022). Approximately 1 in 10 %-infected cats develop mutations that result in feline infectious peritonitis. In large multi-cat situations, FECV is shed in the feces of most apparently healthy cats and transmission occurs through direct contact with feces or contaminated bedding and other fomites (Pedersen et al 2004). Kittens become infected at approximately 9 weeks of age (Pedersen et al. 2008). The time between onset of clinical signs and death also varies, but younger cats and cats with effusive disease have a shorter course of disease than older cats and cats with noneffusive disease (Pedersen 2014). Even with severe FIP, some cats can live for months. In situations where multiple cats are present, feline enteric coronavirus (FECV) is extremely common and highly contagious. Almost all cats that come into contact with FECV from shedding cats will become ill, but otherwise the infection is usually asymptomatic or causes only mild, transient diarrhea (Pedersen et al 2008; Vogel et al 2010; Ermakov et al 2021). On the other hand, feline infectious peritonitis virus (FIPV) is not transmitted by the fecal-oral route but originates from avirulent FECV in a small percentage of infected cats and causes feline infectious peritonitis (FIP) (Pedersen et al 1981; Vennema et al 1998). Anorexia, lethargy, weight loss, pyrexia, ocular and neurological signs such as gait abnormalities or inadequate mentation are nonspecific (Giori et al 2011; Kipar et al 2014). The infection has two forms: “wet” and “dry”. The dry form causes inflammatory changes around the vessels, seizures, ataxia and excessive thirst, while the wet form leads to abdominal distension due to excessive accumulation of fluid in the abdominal cavity. Specificity is always the most important diagnostic value to consider to avoid misdiagnosis of FIP in unaffected cats.
2. Diagnostic tests for feline infectious peritonitis
Diagnosis is based on the cat's age, background, clinical signs, and physical examination. Abdominal distension with ascites, dyspnea with pleural effusion, jaundice, hyperbilirubinuria, palpable masses in the kidneys and/or mesenteric lymph nodes, uveitis, and various neurologic signs associated with brain and/or spinal cord involvement are common in cats with FIP, with retinal changes being the most common ocular involvement. Retinal vascular cuffing may occur, appearing as blurred grayish lines on either side of the vessels. Granulomatous changes in the retina are occasionally seen. FIPV infection has been shown to be associated with T-cell depletion by apoptosis, although the virus cannot infect CD4+ and CD8+ T-cells (Haagmans et al. 1996; De Groot et al. 2005). Given the high mortality rate, many veterinarians and pet owners are cautious about making a diagnosis based on “reasonable certainty.” The challenge is to decide whether a test increases the likelihood that the clinical signs are due to FIP (indirect tests) or offers a definitive diagnosis (direct tests). It is essential to recognize that the sensitivity and specificity of any indirect test will vary depending on how likely the cat is to be infected based on other factors. This means that the positive predictive value of a test such as a complete blood count (CBC) or albumin:globulin (A:G) ratio for predicting FIP will be much higher in cats with FIP-like signaling than in cats with non-FIP signaling. It should be noted that the results of other indirect tests are only estimates, and the results of additional indirect tests have the potential to both confound and aid the diagnostic process.
3. Diagnostic tests
The problem with FIP diagnostics is that non-invasive tests are not reliable enough. In general, effusion tests have a significantly higher predictive value than blood tests (Stranieri et al. 2018; Hartmann et al. 2003). As a result, the identification of FIP ante mortem in cats without significant effusion is particularly difficult. The most useful ante-mortem indicators are positive anti-Corona (IgG) antibody titers in cerebrospinal fluid (CSF), high total serum protein, and MRI changes such as periventricular contrast enhancement, ventricular dilatation, and hydrocephalus. However, monoclonal antibodies from affected tissues and coronavirus-specific polymerase chain reaction (PCR) are valuable in post mortem evaluations (Foley et al. 1998). As a clear diagnosis cannot be made on the basis of symptoms, medical history and clinical and laboratory indicators alone, these factors should always be considered as a whole, sometimes in combination with other factors such as molecular or even more invasive diagnostic procedures.
3.1. Analysis of effusion samples
In cases of suspected FIP with effusion, an effusion sample can be incredibly helpful in establishing the diagnosis and then the hematological findings, so obtaining effusion samples should always be a top priority. In cases of ascites, a sample can be obtained by ultrasound-guided fine needle aspiration or the “flying cat” technique. For identifying small amounts of fluid in the chest and abdomen, ultrasonography provides a useful aid in localizing effusion pockets in the abdomen, while evidence of pericardial effusions can be obtained by muffled cardiac sounds and electrocardiographic changes. Ultrasonography should be used repeatedly to identify any small volume effusions, and ultrasonography can also be used to guide sampling of small pockets of fluid. In cats with pericardial effusions, auscultation of the heart will reveal muffled sounds and the ECG will reveal typical changes. FIP effusions are often clear, viscous / sticky, straw yellow and rich in protein (cytology often describes a dense eosinophilic protein background) with a total protein concentration> 35 g / l (> 50 % globulins). Chlootic effusions are rarely described. FIP effusions are often pyogranulomatous in nature with macrophages, non-degenerate neutrophils and a relatively small number of lymphocytes. As a result, effusions are often referred to as modified transudates based on cell number (<5 × 109 cells / L), but exudates based on protein concentration (greater than 35 g / L). Typical FIP effusions have a low A: G ratio (see above) and an increased AGP content, which is similar to the serum content. A recent study (Hazuchova et al. 2017) found that AGP concentrations in effusions (> 1.55 mg / ml) are more useful (sensitivity and specificity 93 %) in distinguishing FIP cases from cases without FIP than serum AGP levels or other APPs. The rival test is a simple test that can be used to distinguish transudate from exudate in a effusion sample (Barker and Tasker 2020). Positive results simply suggest that the effluent is exudate and not specific to FIP; positive transudate results have been documented in situations other than FIP (eg, bacterial / septic peritonitis and lymphoma) (Fischer et al. 2012).
3.2. Serum biochemistry
Although the changes in blood biochemistry observed in FIP cases are variable and often non-specific, there are several key anomalies that need to be addressed in order to confirm the diagnosis of FIP.
3.2.1. Acute phase proteins
Many inflammatory and non-inflammatory diseases produce acute phase proteins (APPs) in the liver in response to cytokines released by macrophages and monocytes (particularly interleukins 1 and 6 and tumor necrosis factor α). AGP is an abbreviation for α1-acid glycoprotein and its examination may help in the diagnosis of FIP. Although the increase in AGP levels (> 0.48 mg / ml) is not specific for FIP, patients with FIP often have significantly high AGP levels (> 1.5 mg / ml). As a result, the magnitude of the increase may be valuable in helping to diagnose FIP, with higher levels more effectively increasing the suspicion index (Giori et al 2011; Hazuchova et al 2017).
3.2.2. Hyperglobulinemia
In 89% cases, hyperglobulinemia is present; often in association with hypoalbuminemia or low-normal serum albumin levels (observed in 64.5 % cases) (Riemer et al. 2016). Hyperproteinemia may not always occur due to the existence of hypoalbuminemia. The albumin: globulin (A: G) ratio is low in hyperglobulinemia and hypoalbuminemia (low-normal albumin concentration) and this parameter can be used to assess the likelihood of FIP in a particular case.
3.2.3. Hyperbilirubinemia
Hyperbilirubinemia occurs in 21-63 % cases of FIP and is more common in effusive FIP, where alanine aminotransferase (ALT), alkaline phosphatase (ALP) and γ-glutamyltransferase enzymes are commonly high (although these may be slightly increased in FIP cases). FIP is rarely associated with hyperbilirubinemia due to immune-mediated hemolytic anemia (IMHA) (Norris et al. 2012) and cats are often not severely anemic. In the absence of high liver enzyme activity or severe anemia, the presence of hyperbilirubinaemia should suggest FIP (note that sepsis and pancreatitis may cause hyperbilirubinaemia without increased liver enzyme activity). Based on a stepwise evaluation of cats with FIP, it was documented that hyperbilirubinemia was more typically recognized in cats just before death or euthanasia than in the first presentation (Harvey et al. 1996). In addition, higher bilirubin levels were observed in cats just before death or euthanasia than in the first presentation.
3.3. Hematology
In FIP, hematological changes are non-specific; however, there are several abnormalities that need to be checked to confirm the diagnosis. Lymphopenia is the most common change (55 - 77%) in cases, with a recent study (Riemer et al. 2016) revealing lymphopenia in only 49.5 % cases of FIP, with neutrophilia (39 - 57 %), left-sided and mild to severe normocytic, normochromic anemia (37-54 %) (Riemer et al. 2016; Norris et al. 2012). Recently, an association between FIP and microcytosis (with or without anemia) has been discovered. FIP can cause severe IMHA with concomitant regenerative anemia; however, this is an unusual phenomenon.
3.4. Serology
ELISAs, indirect immunofluorescence antibody assays, and rapid immunocompromination assays are the most common assays for anti-FCoV antibodies in serum (Addie et al. 2015). In most studies, cells infected with CoV pigs or cats are used as substrate and titers are measured in multiples of serum dilutions. A positive anti-FCoV antibody test means that the cat has been infected with FCoV and has seroconverted (which lasts 2-3 weeks after infection). The tests are therefore of limited clinical importance. Breed-dependent differences in anti-FCoV antibody titers were found, which could indicate differences in the breed's response to FCoV infection (Meli et al. 2013). Although cats with FIP had higher anti-FCoV antibody titers than cats without FIP, no difference was found between the median anti-FCoV antibody titers in healthy cats and cats with suspected FIP. As a result, the titer in one animal is only marginally useful in identifying cats with FIP (Bell et al. 2006). Many clinically healthy cats (especially those in multi-cat households) have positive and often very high anti-FCoV antibody titers, while 10 % cats with FIP are seronegative, which could be due to virus binding to the antibody and its inaccessibility for serological testing. which also points to problems with interpretation (Meli et al 2013). A negative FCoV antibody test if dry FIP is suspected may be more effective at excluding FIP (Addie et al. 2009). Nevertheless, negative results have been observed in neurological FIP situations (Negrin et al 2007). As a result, doctors disagree on whether to perform a serological test in suspected cases, even though a positive result almost always means FCoV exposure.
3.5. Current trends in diagnostics
The use of anti-coronavirus antibody testing in cerebrospinal fluid (CSF) for diagnosis in cases involving the central nervous system is another breakthrough in which IgG is detected in the CSF. However, in most cases, the antibody was detected only in cats with high serum IgG titers (Boettcher et al. 2007) An important difference between feline coronavirus and FIP infection is the behavior of the NSP3c gene. The infected tissue isolates from the second case were found to have a disrupted gene 3c, while in the first case the gene was intact. Mutation of the S1 / S2 locus and modulation of the furin recognition site, which is normally present in the S-gene of the enteric coronavirus (Levy and Hutsell 2019), is also a crucial contributing factor. The diagnostic utility of cerebrospinal fluid immunocytochemistry is also used to diagnose FIP manifested by severe central nervous system involvement. Immunocytochemical staining (ICC) of feline coronavirus antibodies in cerebrospinal fluid macrophages is a highly sensitive test, especially for the diagnosis of ante mortem with a sensitivity of 85 % and a specificity of 83.3 % (Gruendl et al 2017).
4. Conclusions
In cats with suspected FIP, anamnesis, clinical signs and clinicopathological examinations should be correlated. The dry form is more difficult to diagnose than the wet form. In the wet form, laboratory analysis of the fluid can be performed, such as the Rivalt's test. If the test is negative, the probability of FIP is small, but if the test is positive, additional diagnostic tests should follow to confirm FIP. In FIP, the A: G ratio is low because hyperglobulinemia and hypoalbuminemia (low normal albumin concentration) are present, and this parameter can be used to assess the likelihood of FIP in a particular case. Patients with FIP often have significantly high levels of AGP (α1-acid glycoprotein). When distinguishing FIP cases from non-FIP cases, AGP concentrations in effusions (> 1.55 mg / ml) have a sensitivity and specificity of 93 %.
Conflict of interests
The authors declare that they do not have a conflict of interest.
Financing
No financial support was provided from any institute or other source.
5. Literature
Addie D, Belak S, Boucraut-Baralon C (2009) Feline infectious peritonitis. ABCD guidelines on prevention and management. J Feline Med Surg 11: 594–604.
Addie D, Belák S, Boucraut-Baralon C, Egberink H, Frymus T, Gruffydd-Jones T, Hartmann K, Hosie MJ, Lloret A, Lutz H (2009) Feline infectious peritonitis. ABCD guidelines on prevention and management. Journal of Feline Medicine and Surgery 11: 594–604.
Addie DD, le Poder S, Burr, P (2015) Utility of feline coronavirus antibody tests. J Feline Med Surg 17: 152–162.
Barker E, Tasker S (2020) Update on feline infectious peritonitis. In Practice 42: 372–383.
Bell ET, Malik R, Norris JM (2006) The relationship between the feline coronavirus antibody titre and the age, breed, gender and health status of Australian cats. Aust Vet J 84: 2–7.
Bell ET, Toribio JA, White JD (2006) Seroprevalence study of feline coronavirus in owned and feral cats in Sydney, Australia. Aust Vet J 84: 74–81.
Boettcher IC, Steinberg T, Matiasek CEG, Hartmann K, Fischer A (2007) Use of anti-corona virus antibody testing of cerebrospinal fluid for diagnosis of feline infectious peritonitis involving the central nervous system. J Am Vet Med Assoc 230: 199-205.
De Groot-Mijnes JD, Van Dun JM, Van der Most RG, de Groot RJ (2005) Natural history of a recurrent feline coronavirus infection and the role of cellular immunity in survival and disease, Journal of Virology 79: 1036–1044
Fischer Y, Sauter-Louis C, Hartmann K (2012) Diagnostic accuracy of the Rivalta test for feline infectious peritonitis. Vet Clin Pathol 41: 558–567.
Foley JE, Lapointe JM, Koblik P, Poland A, Pedersen NC (1998) Diagnostic features of clinical neurologic feline infectious peritonitis. J Vet Intern Med 12: 415423.
Giori L, Giordano A, Giudice C (2011) Performances of different diagnostic tests for feline infectious peritonitis in challenging clinical cases. J Small Anim Pract 52: 152–157.
Gruendl S, Matasek K, Matiasek L, Fischer A, Felten S, Jurina K, Hartmann K (2017) Diagnostic utility of cerebrospinal fluid immunocytochemistry for diagnosis of feline infectious peritonitis manifesting in central nervous system. J Feline Med Surg 19: 576–585.
Haagmans BL, Egberink HF, Horzinek MC (1996) Apoptosis and T-cell depletion during feline infectious peritonitis. J Virol 70: 8977–8983
Hartmann K, Binder C, Hirschberger J, Cole D, Reinacher M, Schroo S, Frost J, Egberink H, Lutz H, Hermanns W (2003) Comparison of different tests to diagnose feline infectious peritonitis. J. Vet. Intern. Med.17: 781–790.
Harvey CJ, Lopez JW, Hendrick MJ (1996) An uncommon intestinal manifestation of feline infectious peritonitis: 26 cases (1986-1993). J Am Vet Med Assoc 209: 1117–1120.
Hazuchova K, Held S, Neiger R (2017) Usefulness of acute phase proteins in differentiating between feline infectious peritonitis and other diseases in cats with body cavity effusions. J Feline Med Surg 19: 809–816.
Healey EA, Andre NM, Miller AD, Whitaker GR, Berliner EA (2022). Outbreak of feline infectious peritonitis (FIP) in shelter-housed cats: molecular analysis of the feline coronavirus S1 / S2 cleavage site consistent with a 'circulating virulent – avirulent theory'of FIP pathogenesis. Journal of Feline Medicine and Surgery Open Reports 8: 20551169221074226.
Kipar A, May H, Menger S, Weber M, Leukert W, Reinacher M (2005) Morphological features and development of granulomatous vasculitis in feline infectious peritonitis, Veterinary Pathology 42: 321–330
Kipar A, Meli ML (2014) Feline infectious peritonitis: Still an enigma? Vet. Pathol. 51: 505–526.
Levy JK, Hutsell S (2019) MSD veterinary manual: Feline infectoius peritonitis (FIP). USA: Merck Sharp and Dohme Corp.
Meli M, Kipar A, Müller C, Jenal K, Gönczi EE, Borel N, Gunn-Moore D, Chalmers S, Lin F, Reinacher M, Lutz H (2004) High viral loads despite absence of clinical and pathological findings in experimental cats infected with feline coronavirus (FCoV) type I and in naturally FCoV-infected cats, Journal of Feline Medicine and Surgery 6: 69–81.
Meli ML, Burr P, Decaro N (2013) Samples with high virus load cause a trend toward lower signal in feline coronavirus antibody tests. J Feline Med Surg 15: 295– 299.
Negrin A, Lamb CR, Cappello R (2007) Results of magnetic resonance imaging in 14 cats with meningoencephalitis. J Feline Med Surg 9: 109–116.
Norris JM, Bosward KL, White JD (2012) Clinico-pathological findings associated with feline infectious peritonitis in Sydney, Australia: 42 cases (1990-2002). Aust Vet J 83: 666–673.
Pedersen NC (2014) An update on feline infectious peritonitis: diagnostics and therapeutics. The veterinary journal 201: 133-141.
Pedersen NC (2009) A review of feline infectious peritonitis virus infection: 1963–2008. J. Feline Med. Surg.11: 225–258.
Pedersen NC (1995) An overview of feline enteric coronavirus and infectious peritonitis virus infections. Feline Practice 23: 7–20.
Pedersen NC, Allen CE, Lyons LA (2008) Pathogenesis of feline enteric coronavirus infection. Journal of Feline Medicine and Surgery 10: 529–541.
Pedersen NC, Liu H, Dodd KA, Pesavento PA (2009) Significance of coronavirus mutants in feces and diseased tissues of cats suffering from feline infectious peritonitis. Viruses 1: 166–184
Pedersen NC, Sato R, Foley JE, Poland AM (2004) Common virus infections in cats, before and after being placed in shelters, with emphasis on feline enteric coronavirus. Journal of Feline Medicine and Surgery 6: 83–88.
Pedersen NC, Boyle JF, Floyd K (1981) Infection studies in kittens, using feline infectious peritonitis virus propagated in cell culture. Am. J. Vet. Res.42: 363– 367.
Riemer F, Kuehner KA, Ritz S (2016) Clinical and laboratory features of cats with feline infectious peritonitis - a retrospective study of 231 confirmed cases (2000-2010). J Feline Med Surg 18: 348–356.
Stranieri A, Giordano A, Paltrinieri, S Giudice C, Cannito V, Lauzi S (2018) Comparison of the performance of laboratory tests in the diagnosis of feline infectious peritonitis. J. Vet. Diagn. Investig. 30: 459–463.
Vennema H, Poland A, Foley J, Pedersen NC (1998) Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology 243: 150–157.
Vogel L, Van der Lubben M, teLintelo EG, Bekker, CP Geerts T, Schuijff LS, Grinwis GC, Egberink HF, Rottier PJ (2010) Pathogenic characteristics of persistent feline enteric coronavirus infection in cats. Vet. Res. 41: 71–82.
Weiss RC, Scott FW (1981) Pathogenesis of feline infectious peritonitis: nature and development of viraemia, American Journal of Veterinary Research 4: 382– 390.