

9.7.2020
S.E. Cook, H. Vogel, D. Castillo, M. Olsen, N. Pedersen, B. G. Murphy
Pôvodný článok: A rational approach to identifying effective combined anticoronaviral therapies against feline coronavirus
Abstrakt
Mačací infekčný zápal pobrušnice (FIP), spôsobený genetickým mutantom mačacieho enterického koronavírusu známym ako FIPV, je smrteľným ochorením mačiek, pre ktoré v súčasnosti nie je k dispozícii žiadna vakcína alebo liečba schválená FDA. Šírenie FIPV u postihnutých mačiek vedie k radu klinických príznakov vrátane kavitárnych výpotkov, anorexie, horúčky a lézií pyogranulomatóznej vaskulitídy a perivaskulitídy s alebo bez postihnutia centrálneho nervového systému a/alebo oka. Vznikla kritická potreba účinných a schválených antivírusových terapií proti koronavírusom vrátane FIPV a zoonotických koronavírusov, ako je SARS-CoV-2, pôvodcu COVID-19. Pokiaľ ide o SARS-CoV-2, predbežné dôkazy naznačujú, že môžu existovať potenciálne klinické a patologické spoločné znaky s mačacím koronavírusovým ochorením, vrátane enterického a neurologického postihnutia. Skúmali sme 89 vytipovaných antivírusových zlúčenín a identifikovali sme 25 zlúčenín s antivírusovou aktivitou proti FIPV, ktoré predstavujú rôzne triedy liekov a mechanizmov antivírusového pôsobenia. Na základe úspešných stratégií kombinovanej liečby u ľudských pacientov s infekciou HIV alebo vírusom hepatitídy C sme identifikovali kombinácie liekov zameraných na rôzne fázy životného cyklu FIPV, ktoré vedú k synergickému antivírusovému účinku. Podobne navrhujeme, aby sa kombinovaná antikoravírusová terapia (cACT) s viacerými mechanizmami účinku a penetrácie do všetkých potenciálnych anatomických miest vírusovej infekcie aplikovala na liečbu ďalších koronavírusov, ako je SARS-CoV-2.
Zhrnutie autora
In vitro sme testovali antivírusovú aktivitu proti FIPV u 89 zlúčenín. Antivírusová aktivita týchto zlúčenín spočivala buď v priamom účinku na vírusové proteíny zapojených do replikácie vírusu, alebo v nepriamom inhibičnom účinku na normálne bunkové procesy uzurpované FIPV na podporu replikácie vírusu. Dvadsaťpäť z týchto zlúčenín vykázalo významnú antivírusovú aktivitu. Zistili sme aj to, že určité kombinácie týchto zlúčenín sú účinnejšie ako samotná monoterapia.
Slovník
| Skratka | Anglický výraz | Slovenský preklad |
|---|---|---|
| PI | protease inhibitor | inhibítor proteázy |
| NPI | nucleoside polymerase inhibitor | nukleozidový inhibítor polymerázy |
| NNPI | non-nucleoside polymerase inhibitor | nenukleozidový inhibítor polymerázy |
| CPE | cytopathic effect | cytopatický efekt |
| cACT | combined anti-coronaviral therapy | kombinovaná antikoronavírusová terapia |
| cART | combined anti-retroviral therapy | kombinovaná antiretrovírusová terapia |
| CRFK cells | Crandell-Rees Feline Kidney cells | Crandell-Rees mačacie obličkové bunky |
Úvod
Mačacia infekčná peritonotída (FIP) je vysoko smrteľné ochorenie bez účinnej vakcíny alebo liečby schválenej FDA. Aj keď patogenéza nie je úplne objasnená, všeobecne sa predpokladá, že FIP je výsledkom špecifických mutácií vo vírusovom genóme minimálne patogénneho a všadeprítomného mačacieho enterického koronavírusu (FECV), ktoré vedú k vzniku virulentného vírusu FIP (FIPV) [1–3]. Tieto mutácie FECV vedú k zmene tropizmu vírusom napadnutej hostiteľskej bunky z intestinálnych enterocytov na makrofágy peritoneálneho typu. Produktívna infekcia makrofágov vírusom FIPV, cielená rozsiahla anatomická diseminácia a imunitne sprostredkovaná perivaskulitída vedú k vysoko smrteľnému systémovému zápalovému ochoreniu FIP [4]. V dôsledku vírusovej diseminácie sa FIP môže prejavovať klinickými príznakmi odrážajúcimi zápal na rôznych anatomických miestach, ktoré môžu potenciálne zahŕňať brušnú dutinu a vnútornosti, hrudnú dutinu, centrálny nervový systém a/alebo oči [5–8]. Kvôli vysokej úmrtnosti FIP zostáva devastujúcim vírusovým ochorením mačiek a výzvou pri stanovení presnej etiologickej diagnózy za súčasného nedostatku dostupných a efektívnych možností liečby [7, 9]. Vývoj účinnej vakcíny proti FIP bol komplikovaný úlohou na protilátkách závislého zosilnenia (ADE) v patogenéze choroby FIP, kde sa ukázalo, že prítomnosť neneutralizujúcich antikoronavírusových protilátok zhoršuje ochorenie FIP [10–12] .
Koronavírusy u cicavcov infikujú a spravidla spôsobujú ochorenie črevného traktu alebo dýchacieho systému infikovaných hostiteľov [13]. FIP sa však často prejavuje ako syndróm multisystémového zápalového ochorenia v dôsledku rozsiahleho šírenia makrofágov infikovaných FIPV. Nedávny pandemický výskyt SARS-CoV-2 má u infikovaných ľudských pacientov za následok rôzne syndrómy choroby, súhrnne označovanej ako COVID-19. Aj keď má SARS-CoV-2 zjavný tropizmus pre respiračný epitel, ktorý vedie k intersticiálnej pneumónii, nedávne dôkazy naznačujú, že COVID-19 sa môže prejavovať aj ako zažívacie ochorenie a klinicky sa prejavovať ako hnačka [14, 15]. Tropizmus pre tieto tkanivá odráža membránovú expresiu proteínu ACE2, bunkového cieľa SARS-CoV-2 [16]. Ďalej sa ukazuje, že SARS-CoV-2 je schopný infikovať a spôsobiť zápalové ochorenie v tkanivách mimo črevného traktu a dýchacích ciest, vrátane mozgu, očí, reprodukčných orgánov a srdcového myokardu [17–21]. Neuroinvazia mozgového kmeňa a následná encefalitída spôsobená SARS CoV-2 môžu prispieť k respiračnému zlyhaniu u pacientov s COVID-19 [20,22]. Experimentálne je SARS CoV-2 schopný vytvoriť produktívnu infekciu u mačiek [23]. Infekcia FIPV mačiek a infekcia SARS CoV-2 u ľudských pacientov sa preto podobajú viac, ako sa pôvodne zdalo.
Vznikla okamžitá a kritická potreba dostupných a účinných antivírusových terapií na liečenie týchto koronavírusových chorôb. Mačky infikované FIPV by mohli slúžiť ako translačný model a poskytnúť užitočné poznatky použiteľné pre pacientov infikovaných SARS-CoV-2 s COVID-19. Posledné antivírusové klinické štúdie u mačiek experimentálne aj prirodzene infikovaných FIPV priniesli nádej pre liečbu a vyliečenie FIP pomocou GS-441524, nukleozidového analógu a metabolitu proliečiva Remdesivir (Gilead Sciences) alebo GC-376, 3C-like inhibítora proteázy FIPV (Anivive) [24–26]. Remdesivir, prodrug GS-441524, sa nedávno ukázal byť sľubným pri liečbe ľudských pacientov infikovaných SARS-CoV-2 [27,28]. Napriek týmto nedávnym klinickým úspechom musia byť tieto antivírusové zlúčeniny ešte schválené a v súčasnosti nie sú k dispozícii na klinické veterinárne použitie u mačiek s FIP.
Identifikácia a vývoj účinných antivírusových terapií môžu byť nákladné aj časovo náročné. Cielený skríning a opakované použitie liekov už schválených FDA alebo schválených na výskumné použitie, môžu hrať efektívnu úlohu pri objavovaní liekov. Použitím predpokladaných antivírusových zlúčenín vybraných na základe ich preukázanej účinnosti pri liečbe iných RNA-vírusov sme identifikovali podskupinu zlúčenín so silnou anti-FIPV aktivitou a charakterizovali ich profily bezpečnosti a účinnosti in vitro. Na základe veľkého úspechu kombinovanej antiretrovírusovej terapie (cART) proti HIV-1 a kombinovanej liečby vírusu hepatitídy C [29] sme vyvinuli metódy na identifikáciu účinných kombinačných terapií proti FIPV. Počiatočné monoterapie proti HIV-1, ako je azidotymidín (AZT), často viedli k vírusovým únikovým mutáciám. Zdá sa, že súčasné použitie viacerých antivírusových zlúčenín blokuje tento adaptívny vírusový evolučný mechanizmus, pretože vývoj HIV-1 je modernými cART účinne zastavený [30]. Úspešnosť cART je výsledkom farmakologického zamerania na viac stupňov životného cyklu vírusu súčasne, pričom sa spoločne dosahuje synergický antivírusový účinok [31].
Vzhľadom na pôsobivý úspech cART by sa mohlo javiť, že súčasné zacielenie na FIPV v rôznych fázach životného cyklu vírusu kombinovanou antikoronavírusovou terapiou (cACT) môže ponúknuť vyššiu úroveň trvalého a úplnejšieho úspechu, v porovaní s monoterapiami samotnými. Zahrnutie antivírusového agenta do cACT schopného preniknúť hematoencefalickou (BBB) a hematookulárnou bariérou, a dosiahnuť farmakologicky relevantné tkanivové koncentrácie môže uľahčiť eradikáciu FIPV v celom systéme. Popisujeme tu súbor in vitro testov uľahčujúcich rýchly skríning a identifikáciu účinných antikoronavírusových zlúčenín. Účinné antivírusové látky s rôznymi mechanizmami účinku a predpokladanou distribúciou v tele boli kombinované do cACT a testované na synergiu zlúčenín. Predpokladali sme, že kombinované použitie dvoch alebo viacerých účinných antivírusových monoterapií s rôznymi mechanizmami účinku uľahčí identifikáciu synergických kombinácií poskytujúcich vynikajúcu antikoronavírusovú účinnosť v porovnaní s ich samostatným použitím. Identifikácia úspešnej cACT môže tiež poskytnúť vodítka k liečbe ďalších objavujúcich sa vírusových ochorení, ako je SARS-CoV-2.
Výsledky
Testovanie zlúčenín
Na identifikáciu zlúčenín s anti-FIPV aktivitou bola in vitro testovaná skupina 89 zlúčenín (doplnková tabuľka 1) z rôznych tried liečiv a s rôznymi predpokladanými mechanizmami účinku. Testované zlúčeniny zahŕňali inhibítory nukleozidovej polymerázy (NPI), nenukleozidové polymerázové inhibítory (NNPI), inhibítory proteázy (PI), NS5A inhibítory, súbor nových anti-helikázových chemických „fragmentov“ a súbor zlúčenín s neurčenými mechanizmami účinku . Z tejto skupiny 89 zlúčenín sa ukázalo, že celkovo 25 rôznych zlúčenín vykazuje antivírusovú aktivitu proti FIPV vrátane NPI, PI, inhibítorov NS5A a dvoch zlúčenín s neurčenými mechanizmami účinku (označovaných ako „iné“, obr. 1). Medzi tieto úspešné antivírusové látky patrili toremiféncitrát, daklatasvir, elbasvir, lopinavir, ritonavir, nelfinavirmesilát, K777 / K11777, grazoprevir, amodiaquín, EIDD 1931, EIDD 2801 a GS-441524 pochádzajúce od troch rôznych čínskych výrobcov (tabuľka 1, obr. 2). Testovali sme niekoľko nukleozidových analógových zlúčenín poskytnutých spoločnosťou Gilead Sciences štrukturálne príbuzných nukleozidovým analógom GS-441524 a Remdesivir na ich antivírusové vlastnosti a našli sme niekoľko s potenciálom (obsiahnutých vo vyššie uvedených 25 identifikovaných zlúčeninách), ale tieto látky sme ďalej nesledovali. Pre ďalšie analýzy bolo vybraných 13 antivírusových látok. Tento celkový počet zahrňuje už pretým identifikovaný 3-C inhibítor proteázy, GC-376 (Anivive).
| Názov zlúčeniny | Kategória lieku | EC50 (µM) |
|---|---|---|
| GC376 | PI | 0.04 |
| EIDD 1931 | NPI | 0.09 |
| Elbasvir | NS5A Inhibitor | 0.16 |
| EIDD 2801 | NPI | 0.4 |
| GS-441524+ | NPI | 0.66 |
| K777/K11777 | PI | 0.67 |
| Toremifene citrate | Iné* | 5 |
| Amodiaquine | Iné** | 6.5 |
| Lopinavir | PI | 8.08 |
| Ritonavir | PI | 8.7 |
| Grazoprevir | PI | 12.13 |
| Nelfinavir mesylate | PI | 13.47 |
EC50 zlúčenín s anti-FIPV aktivitou.
PI = Inhibítor proteázy; NPI = Nukleozidový inhibítor polymerázy
+ MedChem Express, HY-103586
* Selektívny modulátor estrogénových receptorov
** 4-Aminoquinoline

Testované zlúčeniny podľa mechanizmu účinku
(A) Všetky testované zlúčeniny
(B) Zlúčeniny, u ktorých bola pri testoch in vitro zistená anti-FIPV aktivita

Príklad testovacej matrice využívajúcej farbenie kryštálovou violeťou na identifikáciu anti-FIPV aktivity pri 10μM.
Horný ľavý riadok sú kontrolné jamky iba s CRFK bunkami a bez liečiva alebo FIPV. Pravý horný riadok je pozitívna kontrola využívajúca GS-441524 so známou úplnou ochranou CRFK buniek pred FIPV-indukovanou bunkovou smrťou. Celý spodný rad jamiek predstavuje CRFK bunky infikované FIPV a bez liečby. Zvyšné riadky sú testovacie jamky, pričom ľavá polovica hodnotí cytotoxicitu pri 10μM (bez infekcie FIPV) a pravá polovica hodnotí anti-FIPV aktivitu pri 10μM pre ktorúkoľvek danú zlúčeninu. Strata zafarbenia naznačuje stratu buniek. Daklatasvir a velpatasvir demonštrovali anti-FIPV aktivitu, o čom svedčí zvýšené farbenie kryštálovou violeťou (relatívne intaktné bunkové monovrstvy) v porovnaní s kontrolnými jamkami obsahujúcimi iba FIPV (spodný rad doštičky). Lieky 32, 33 a 34 vykazovali neprítomný až minimálny antivírusový účinok, zatiaľ čo liečivo 34 (Ravidasvir) tiež vykazovalo cytotoxicitu pri koncentrácii 10μM na základe dramatického vyčistenia jamiek pozorovaného v ľavej polovici matrice bez FIPV.
Stanovenie antivírusovej účinnosti
Antivírusová účinnosť (EC50) sa stanovila pre 10 antivírusových zlúčenín. Pre tieto zlúčeniny bola EC50 v rozmedzí od 0,04μM do 13,47μM (tabuľka 1, obr. 3). Jedna z antivírusových látok, Daclatasvir, preukázal neprijateľnú cytotoxicitu pri 20μM a bol vyradený z ďalšieho testovania. Ukázalo sa, že GS-441524 pochádzajúci z Číny (MedChemExpress, HY-103586) má porovnateľnú EC50 v porovnaní s predtým publikovanými hodnotami pre GS-441524 pochádzajúci z Gilead Sciences [25].
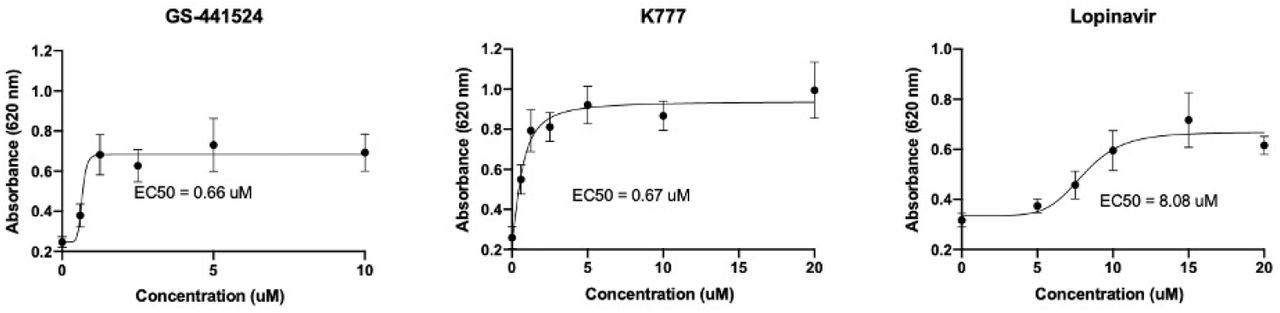
Reprezentatívne príklady nelineárnych regresných analýz EC50 pre zlúčeniny s anti-FIPV aktivitou.
Na identifikáciu polovičnej maximálnej efektívnej koncentrácie (EC50) sa uskutočnili sériové riedenia každej zlúčeniny s anti-FIPV aktivitou. Tu uvedené výsledky GS-441524 predstavujú zlúčeninu pochádzajúcu z MedChemExpress.
Bezpečnostné profily cytotoxicity
Profily bezpečnosti na cytotoxicitu (CSP) sa stanovili pre desať rôznych antivírusových zlúčenín v bunkách CRFK. Pri 5μM sedem testovaných zlúčenín nevykazovalo v podstate žiadnu cytotoxicitu, zatiaľ čo dve z antivirotík, amodiaquine a toremifene, mali 11, respektíve 12% cytotoxicitu (obr. 4; tabuľka 2). 50% cytotoxická koncentrácia (CC50) pre GC376 sa uvádza ako >150μM [32]. Je zaujímavé, že na základe testu Promega CellTox ™ Green Cytotoxicity bola cytotoxicita obidvoch EIDD zlúčenín v podstate nedetegovateľná až do 100μM. Vizuálna kontrola jamiek s EIDD tesne pred aplikáciou fluorescenčného farbiva a odčítaním z matrice však odhalila rozdiely v morfológii buniek (cytopatický účinok) medzi neliečenými CRFK bunkami a liečenými bunkami. Neliečené CRFK bunky sa vyznačovali adherentnou vretenovou morfológiou v jednej monovrstve, zatiaľ čo jamky s EIDD preukázali zreteľný pokles konfluencie v porovnaní s variabilnou morfológiou buniek vrátane zaokrúhľovania buniek (cytopatický účinok). Rozpor medzi subjektívnym vizuálnym hodnotením jamiek s EIDD a fluorescenčným testom je záhadou. Je možné, že celkové zníženie počtu buniek v jamkách s EIDD viedlo k strate a degradácii nukleovej kyseliny nevyhnutnej na fluorescenčné viazanie a detekciu v teste CellTox.
| Zlúčenina | Kategória lieku | 5μM | 10μM | 25μM | 50μM | 100μM |
|---|---|---|---|---|---|---|
| Elbasvir | NS5A Inhibítor | 0.67 | 0.46 | 1.4 | 2 | 4.9 |
| K777/K11777 | PI | 0.61 | 0.29 | 2.3 | 9 | 16 |
| Toremifene citrate | Iná* | 12 | 22 | 23 | 35 | 39 |
| Amodiaquine | Iná** | 11 | 12 | 19 | 23 | 26 |
| Lopinavir | PI | 0.67 | 15 | 13 | 16 | 18 |
| Ritonavir | PI | 0.49 | 0.84 | 2 | 33 | 26 |
| Grazoprevir | PI | 0.55 | 0 | 0.81 | 3.9 | 19 |
| EIDD1931 | NPI | 1.2 | 0.94 | 0.6 | 0.78 | 2.8 |
| EIDD2801 | NPI | 0 | 0.95 | 0.63 | 1.7 | 0.8 |
| GS-441524+ | NPI | 0.21 | 0 | 0 | 0 | 0 |
Percento cytotoxicity podľa zlúčeniny a koncentrácie.
PI = Inhibítor proteázy; NPI = Nukleozidový inhibítor polymerázy
* Selektívny modulátor estrogénových receptorov
** 4-Aminoquinoline
+ NMPharmTech

Reprezentatívne profily cytotoxicity.
Stĺpcové grafy percenta cytotoxicity +/- štandardná odchýlka (SD) pre štyri zlúčeniny s anti-FIPV aktivitou. Percentuálne hodnoty cytotoxicity sa stanovili normalizáciou cytotoxicity pre kontrolné jamky s pozitívnou toxicitou (nastavená na 100% cytotoxicitu) a neliečené bunky CRFK (nastavené na 0% východiskovej cytotoxicity).
Kvantifikácia inhibície produkcie vírusovej RNA pri monoterapii
Na meranie schopnosti každej antivírusovej látky inhibovať koronavírusovú replikáciu pri monoterapii (Vírusový RNA knock-down test) sa použil test RT PCR v reálnom čase. Zlúčeninami demonštrujúcimi najväčšiu inhibíciu produkcie FIPV RNA boli GC376, 3C-like inhibítor koronavírusovej proteázy, GS-441524, EIDD-1931 a EIDD-2801, pričom posledné tri boli nukleozidové analógy (obr. 5, tabuľka 3). Medzi látky s najmenším inhibičným účinkom na produkciu vírusovej RNA patria elbasvir, nelfinavir a ritonavir. Ritonavir, inhibítor proteázy, sa používa v kombinácii s lopinavirom na liečbu infekcie HIV-1 (Kaletra, AbbVie). Monoterapia lopinavirom má u ľudí neuspokojivú orálnu biologickú dostupnosť, avšak pri použití v kombinácii sa ukázalo, že ritonavir výrazne zlepšuje plazmatickú koncentráciu lopinaviru [33]. Preto aj napriek relatívne minimálnej inhibícii FIPV identifikovanej s ritonavirom ako monoterapiou, bola táto zlúčenina predmetom ďalšieho testovania vrátane kombinovaného antikoravírusového hodnotenia.
| Zlúčenina | Násobok redukcie titra vírusu |
|---|---|
| GC376 (20μM) | 25000 |
| GC376 | 7300 |
| GS-441524 (NMPharmTech) | 5280 |
| EIDD-1931 | 3700 |
| GS-441524 (MedChemExpress) | 3500 |
| EIDD-2801 | 2110 |
| Lopinavir | 309 |
| Toremifene | 10 |
| K777 | 7 |
| Grazoprevir | 5 |
| Amodiaquine | 4 |
| Elbasvir | 2 |
| Nelfinavirmesylate | 1 |
| Ritonavir | 1 |
Násobné zníženie počtu kópií vírusovej RNA pre anti-FIPV zlúčeniny pri monoterapii
*Pokiaľ nie je uvedené inak, všetky zlúčeniny sa použili pri 10 μM.

Násobné zníženie počtu kópií FIPV RNA s použitím antivírusových zlúčenín pri monoterapii.
FIPV-infikované CRFK bunky sa inkubovali 24 hodín so zlúčeninami so zistenou anti-FIPV aktivitou. Počet vírusových kópií sa následne stanovil pomocou RT-qPCR a normalizoval sa na počet mačacích GAPDH kópií, aby sa určil účinok násobného zníženia pre každú zlúčeninu. Všetky zlúčeniny sa testovali pri 10μM, pokiaľ nie je uvedené inak. Všetky experimentálne ošetrenia sa uskutočňovali v triplikátových jamkách a násobný pokles sa vypočítal vydelením priemerného experimentálneho normalizovaného počtu kópií FIPV priemerným normalizovaným počtom kópií FIPV stanoveným pre neošetrené jamky infikované FIPV.
1GS-441524 – NMPharmTech (Čína).
2GS-441524 – MedChemExpress (Čína).
Kvantifikácia inhibície produkcie vírusovej RNA pri cACT
Na identifikáciu kombinácií liekov so synergickým antivírusovým účinkom oproti monoterapii sa vybrali kombinácie dvoch alebo viacerých zlúčenín na základe (i) zavedených kombinácií používaných pri iných vírusových infekciách, ako je HIV-1 a HCV, (ii) liekov s rôznymi mechanizmami účinku , (iii) potenciálnych zmien v systémovej distribúcii zlúčeniny (napr. schopnosti preniknúť cez hematoencefalickú alebo hematookulárnu bariéru podľa chemickej klasifikácie) a (iv) minimálnej cytotoxicity (na základe CSP). Pre každú cACT bolo akékoľvek výsledné zníženie počtu kópií FIPV prevyšujúce vypočítaný aditívny účinok pre každé liečivo použité v režime monoterapie považované za synergické (tabuľka 4). Najväčší synergický účinok s najvyšším celkovým násobným znížením vírusovej RNA, dosiahla kombinácia GC376 a amodiaquinu so 76-násobným znížením vírusovej RNA v porovnaní s aditívnym účinkom (obr. 6). Táto konkrétna synergická kombinácia bola jedným z prekvapujúcich výsledkov vzhľadom na to, že samotný amodiaquín preukázal iba obmedzenú inhibíciu kópií vírusovej RNA FIPV stanovenú pomocou qRT-PCR.
| Zlúčenina 1 | Zlúčenina 2 | Zlúčenina 3 | Redukcia titra vírusu | Add | cACT/add |
|---|---|---|---|---|---|
| GC376 (20 uM) | Amodiaquine | — | 1897000 | 25004 | 76 |
| GC376 (20 uM) | Amodiaquine | Toremifene | 256000 | 25014 | 10 |
| GC376 (20 uM) | K777 | — | 248000 | 25007 | 10 |
| GC376 (20 uM) | Toremifene | — | 128000 | 25010 | 5.1 |
| GC376 (20 uM) | Nelfinavir mesylate | — | 91100 | 25001 | 3.6 |
| Elbasvir (5 uM) | Lopinavir | — | 14000 | 311 | 45 |
| Elbasvir (5 uM) | GC376 (20 uM) | — | 12600 | 25002 | 0.50 |
| GC376 (10 uM) | Amodiaquine | — | 11700 | 7304 | 1.6 |
| GC376 (10 uM) | Grazoprevir | — | 8290 | 7305 | 1.1 |
| GC376 (20 uM) | GS-441524 (NMPharmTech) | 8260 | 30280 | 0.27 | |
| GC376 (10 uM) | Amodiaquine | Elbasvir (5 uM) | 7570 | 7306 | 1.0 |
| GC376 (10 uM) | GS-441524 (MedChem) | — | 6910 | 10800 | 0.64 |
| GC376 (20 uM) | Ritonavir | — | 6560 | 25001 | 0.26 |
| K777 | Lopinavir | — | 5530 | 316 | 18 |
| GC376 (10 uM) | GS-441524 (NMPharmTech) | — | 4340 | 12580 | 0.34 |
| GC376 (20 uM) | Lopinavir | — | 3400 | 25309 | 0.13 |
| Lopinavir | Ritonavir | — | 3130 | 310 | 10 |
| Lopinavir | Ritonavir | Toremifene | 1740 | 320 | 5.4 |
| GC376 (10 uM) | EIDD-2801 | — | 255 | 9410 | 0.03 |
| K777 | Toremifene | — | 25 | 17 | 1.5 |
Násobné zníženie počtu kópií FIPV vírusovej RNA pri kombinovanej terapii (cACT).
Očakávaný aditívny účinok reflektuje súčet násobného zníženia vírusovej RNA každej zlúčeniny použitej pri monoterapii (Tabuľka 3)
* Pokiaľ nie je uvedené inak, všetky zlúčeniny boli použité pri 10μM.
cACT/add – pomer redukcie titra FIPV pri kombinovanej terapii oproti súčtu násobkov redukcie titra pri monoterapii
Add – súčet násobkov redukcie titra pri monoterapii

Vybrané príklady násobného zníženia počtu kópií FIPV RNA pomocou kombinovanej terapie (cACT).
Stĺpce predstavujú priemerný násobný pokles troch jamiek liečených buniek CRFK v porovnaní so stredným násobným poklesom troch neliečených jamiek infikovaných FIPV. Všetky zlúčeniny boli testované pri 10μM, pokiaľ nie je uvedené inak.
Vzhľadom na výraznú anti-FIPV aktivitu GC-376, ako aj na jeho potenciálnu dostupnosť pre pokrok vo farmakokinetických štúdiách in vivo, klinických testoch a nádejnom použití v aplikácii cACT bola táto zlúčenina vybraná na sériu „vírusových RNA knock-down“ testov pri mono a kombinovanej terapii (obr. 7). Celkovo GC376 preukázal vynikajúcu anti-FIPV aktivitu pri 20μM tak pri monoterapii, ako aj pri kombinovanej terapii in vitro. K najvýraznejšiemu zníženiu FIPV RNA došlo v kombinácii GC376 pri 20μM s amodiaquinom pri 10μM. Pokus kombinujúci GC376 s amodiaquinom sa opakoval a oba výsledky sú uvedené pre porovnanie na obr. 7C.

GC376 antivírusová aktivita pri monoterapii a v kombinovanej terapii pri 10 a 20 μM
(A) Redukcia FIPV RNA kvantifikovaná pomocou RT-qPCR s použitím GC376 ako monoterapii pri 10 a 20 μM. Medzi týmito dvoma koncentráciami je významný rozdiel, pričom 20μM je lepšia ako 10μM. (nepárový t-test; p <0,0001).
(B) Kombinovaná in vitro terapia s použitím GC376 pri 10 μM.
(C) Kombinovaná in vitro terapia proti FIPV s použitím GC376 pri 20 μM.
Diskusia
Pretože v súčasnosti neexistuje účinná vakcína proti FIP, existuje silná klinická a celosvetová potreba účinných možností antivírusovej liečby pre mačky infikované FIPV. Testovali sme 89 zlúčenín, ktorých výsledkom je identifikácia 25 antivírusových látok s antivírusovou účinnosťou a silnými bezpečnostnými profilmi proti mačaciemu koronavírusu, FIPV. Identifikovali sme tiež kombinácie antivírusových látok (cACT), ktoré viedli k vyššej účinnosti alebo synergizmu oproti samotnej monoterapii. Obzvlášť zaujímavé bolo zistenie týkajúce sa použitia elbasviru, ktorý opakovane demonštroval vynikajúcu ochranu CRFK pred FIPV vyvolanom CPE pri koncentrácii nižšej ako 1 μM na základe viacerých testov (EC50 0,16 μM). V zásade sa však nezistil žiadny rozdiel v počte kópií vírusovej RNA medzi infikovanými bunkami liečenými elbasvirom alebo bez neho. Ďalšia vizuálna analýza buniek CRFK infikovaných FIPV, ktoré boli liečené elbasvirom, odhalila atypickú morfológiu buniek vo vzťahu k neinfikovaným bunkám, ktorá sa vyznačovala premenlivým zväčšením, zaoblením buniek a čiastočným odlúčením buniek (cytopatický účinok). Tieto „atypické bunky“ sa zriedka oddelili od kultivačnej doštičky a vo výsledku boli hodnoty absorbancie porovnateľné s neinfikovanými kontrolnými jamkami. Tento dichotomický výsledok medzi analýzou doštičky a vírusovým RNA knock-down testom naznačuje, že k antivírusovému účinku elbasviru dochádza po vírusovej replikácii a vo výsledku nemusí elbasvir chrániť bunky pred akumuláciou vírusovej RNA. Elbasvir sa používa na liečbu pacientov infikovaných vírusom hepatitídy C (HCV) a predpokladá sa, že je cielený na HCV NS5A proteín zabraňujúci replikácii a tiež na kompletizáciu viriónov [34]. Aj keď nie je pre FIPV identifikovaný homológ NS5A, je možné, že elbasvir vykazuje podobný antivírusový účinok tým, že zabraňuje kompletizácii viriónov FIPV bez blokovania syntézy vírusovej RNA v CRFK bunkách. Dodatočné vyhodnotenie liečených buniek CRFK infikovaných FIPV pomocou transmisnej elektrónovej mikroskopie môže objasniť účinok elbasviru na ochranu CRFK pred poškodením a smrťou spojenou s FIPV.
Ukázalo sa, že súčasné podávanie ritonaviru s lopinavirom významne zvyšuje plazmatické koncentrácie lopinaviru u potkanov, psov a ľudí [33]. Ritonavir je silný inhibítor CYP3A, ktorý je primárnym enzýmom zodpovedným za metabolizmus inhibítorov proteázy, a preto jeho súbežné podávanie s inými inhibítormi proteázy vedie k zvýšeným systémovým koncentráciám súčasne podávaného inhibítora proteázy, ako je lopinavir [38, 39]. Zvýšenie antivírusového účinku lopinaviru spojené s ritonavirom bolo v testoch eliminácie vírusovej RNA relatívne minimálne s iba 10-násobnou inhibíciou FIPV oproti aditívnemu účinku. Môže to byť výsledkom in vitro artefaktu testovania na mačacej obličkovej bunkovej línii (tj. CRFK bunkách), v ktorej chýba enzým CYP3, čo je enzým, ktorý sa typicky vyskytuje v miestach s vysokým metabolizmom inhibítora proteázy pri efekte prvého priechodu (tj. v enterocytoch a hepatocytoch) [39]. Tieto výsledky naznačujú, že samotné testy in vitro nemusia úplne predpovedať účinok antivírusových látok u mačiek infikovaných FIPV in vivo.
Grazoprevir, inhibítor serínovej proteázy NS3/4, sa používal v kombinácii s elbasvirom, inhibítorom NS5A, na liečbu pacientov infikovaných HCV (Zepatier, Merck) [40]. Tu sme preukázali, že grazoprevir má anti-FIPV aktivitu, keď sa používa ako monoterapia. Inhibítor cysteínovej proteázy K777/K11777 bol skúmaný kvôli svojej schopnosti blokovať vstup koronavírusu (MERS-CoV a SARS-CoV-1) a ebolavírusu a bolo zistené, že úplne inhibuje koronavírusovú infekciu, ale iba v cieľových bunkových líniách bez vírus aktivujúcich serínových proteáz. [41]. V prípade iných bunkových línií K777 inhiboval vstup koronavírusových buniek v kombinácii s inhibítorom serínovej proteázy [41]. Je možné, že obmedzená inhibícia produkcie FIPV RNA K777 by sa mohla zvýšiť, ak by sa kombinovala s inhibítorom serínovej proteázy.
Amodiaquine je antimalarické liečivo a patrí do triedy 4-aminochinolínových liečiv. Amodiaquine spolu s príbuznými 4-aminochinolínmi, ako sú chlorochin a hydroxychlorochin, bol pôvodne vyvinuý pre liečbu malárie [42], a podobne ako chlorochín a hydroxychlorochín oplýva rozsiahlym anatomickým distribučným objemom vrátane očí a mozgu [43–49]. Prenikanie antivírusových látok do CNS a/alebo očných kompartmentov je obzvlášť dôležité v prípade mačiek infikovaných FIPV s neurologickým a/alebo očným postihnutím. Zatiaľ čo niekoľko výskumov definovalo antivírusové vlastnosti chlorochínu a hydroxychlorochínu [28, 50, 51], antivírusová aktivita amodiaquinu sa skúmala aj s identifikáciou antivírusovej aktivity proti vírusu dengue, vírusu Ebola a ťažkej horúčke s vírusovým trombocytopénnym syndrómom (SFTS) [52–55]. Mechanizmus účinku amodiaquinu môže zahŕňať zvýšenie cytoplazmatického lyzozomálneho a/alebo endozomálneho pH, ktoré zabráni uvoľneniu životaschopných viriónov do cytoplazmy [56]. Vzhľadom na svoj jedinečný status triedy liečív a predpokladanú schopnosť prekonať hematoencefalickú bariéru [57] je amodiaquine sľubným kandidátom pre kombinovanú liečbu neurologických a/alebo okulárnych foriem FIP.
Toremiféncitrát, selektívny modulátor estrogénových receptorov (SERM), sa používa na liečbu metastatického karcinómu prsníka u ľudských pacientov. V poslednej dobe bol toremifén hodnotený z hľadiska jeho antivírusových vlastností a preukázal antikoravírusovú aktivitu proti zoonotickým koronavírusom, koronavírusom blízkovýchodného respiračného syndrómu (MERS-CoV) a SARS-CoV-1 [58]. Toremifén tiež preukázal aktivitu proti vírusu Ebola (EBOV) [59, 60]. Aj keď presný mechanizmus antivírusového pôsobenia nie je definovaný, zdá sa, že antivírusový účinok toremifénu proti EBOV spočíva v destabilizácii glykoproteínu EBOV [59].
Je zaujímavé, že GC376 preukázal pri kombinovanej terapii mätúce rozdiely medzi dávkovaním 10 μM a 20 μM . Pri kombinovanom použití pri 10 μM s inými zlúčeninami absentoval synergizmus a v niektorých prípadoch pokles antivírusového účinku v porovnaní s adititívnymi hodnotami v rozmedzí od 0,03 do 1,6 (tabuľka 4). Ak sa použil pri 20 μM, stále existovali prípady, keď kombinácia s inou zlúčeninou viedla k zníženému antivírusovému účinku v porovnaní s GC376 použitým ako monoterapia pri 20 μM. Oveľa viac variácií však pri kombináciách pri 20 μM vykazovalo násobok účinku oproti aditívnym hodnotám v rozmedzí od 0,13 do 76 (tabuľka 4). Konkrétnym príkladom je kontrast medzi GC376 pri 20 μM v porovnaní s GC376 pri 10 μM kombinovanom s amodiaquinom pri 10 μM. Prvý spôsobil najväčšiu inhibíciu vírusovej RNA, ako aj najväčší násobok aditívneho (synergického) účinku, zatiaľ čo druhý spôsobil takmer stratu synergizmu s hodnotou násobku aditívnej hodnoty 1,6.
Identifikácia účinných antivírusových stratégií pre liečbu mačiek infikovaných FIPV má translačné dôsledky pre prebiehajúcu pandémiu SARS-CoV-2. Infekcia FIPV u mačiek pripomína infekciu koronavírusmi u fretiek [61, 62] a porovnáva sa s patogenézou iných chronických chorôb závislých od makrofágov, ako je tuberkulóza [63]. Pretože sa klinické a patogénne podrobnosti infekcie SARS-CoV-2 u ľudí stále objavujú, zdá sa, že existuje určité prekrytie s FIPV v anatomickej distribúcii, klinických prejavoch a pravdepodobnej reakcii na určité antivírusové terapie. U mačiek je mačací enterický koronavírusový biotyp (FECV) obmedzený na zažívací trakt v dôsledku tropizmu enterocytov. Klinické príznaky u mačiek infikovaných FECV siahajú od mierneho gastrointestinálneho ochorenia (hnačka) až po absenciu príznakov. Mutovaný mačací biotyp koronavírusu FIPV získava makrofágový tropizmus a prednostne sa zameriava na serózne povrchy brušnej a hrudnej dutiny s podskupinou mačiek, ktoré demonštrujú postihnutie CNS alebo očí [7]. Podobne u pacientov s COVID-19 existujú správy o hnačkách a podskupine pacientov s postihnutím CNS [14]. Aj keď bol bunkový receptor pre SARS-CoV-2 identifikovaný ako ACE2 [64], bunkový receptor pre FIPV sérotypu I sa ešte musí určiť. Bunkový receptor pre menej klinicky relevantný FIPV sérotypu II bol identifikovaný ako mačacia aminopeptidázová peptidáza (fAPN) [4]. Štúdia využívajúca RNAseq na hodnotenie profilov génovej expresie buniek ascitu získaných z mačiek s FIP neidentifikovala expresiu ACE2, čo naznačuje, že ACE2 pravdepodobne nebude receptorom FIPV sérotypu I [63]. Je potrebné podrobnejšie preskúmanie identity receptora FIPV sérotypu I.
Doterajšie klinické úspechy s použitím GS-441524 alebo GC-376 u mačiek s experimentálnymi a prirodzene sa vyskytujúcimi FIP ukazujú, že je možné účinne liečiť FIP, avšak liečba suchých (granulomatóznych), neurologických a okulárnych FIP stále predstavuje výzvu. Inhibítor proteázy 3C-like proteázy, GC-376, sa javí ako relatívne účinný pri liečbe efúznej infekcie FIPV obmedzenej na telesné dutiny, ale môže byť menej účinný pri liečbe neurologických alebo okulárnych foriem ochorenia [24]. Tieto odlišné výsledky môžu byť výsledkom neúčinnej penetrácie hematoencefalickej a hematookulárnej bariéry, čo robí z GC-376 nádejného kandidáta na kombinovanú terapiu s antivírusovým liekom prenikajúcim do CNS.
Materiály a metódy
Inokulácia FIPV pre in vitro experimenty
Mačacie obličkové bunky Crandell-Reese (CRFK, ATCC) sa kultivovali v bankách T150 (Corning), naočkovali sa FIPV sérotypu II (WSU-79-1146, GenBank DQ010921) a množili sa v 50 ml Dulbeccovho modifikovaného Eagle média (DMEM) s 4,5 g/l glukózy (Corning) a 10% fetálneho hovädzieho séra (Gemini Biotec). Po 72 hodinách inkubácie pri 37°C bol zaznamenaný rozsiahly cytopatický účinok (CPE) a veľké plochy clearingu/oddeľovania buniek. Banky sa následne bleskovo zmrazili na -70°C počas 8 minút, krátko sa rozmrazili pri izbovej teplote a bunky a supernatant sa potom centrifugovali pri 1500 g počas 5 minút, po ktorých nasledoval druhý krok centrifugácie pri 4000 g počas 5 minút, aby sa izolovali bezbunkové vírusové objemy. Supernatant obsahujúci vírusový základ sa rozdelil na 0,5 a 1,0 ml alikvotné časti v 1,5 ml kryotubusoch (Nalgene) a archivoval sa pri -70°C. Po zmrazení sa jedna skúmavka nechala rozmrznúť a titer vírusu sa stanovil pomocou biologických testov (TCID50) a RT PCR metód v reálnom čase (nižšie).
Infekčná dávka dose-50 pre tkanivovú kultúru (TCID50) sa stanovila pomocou vírusového plaque testu. CRFK bunky sa pestovali na 96-jamkovej doštičke pre tkanivové kultúry (Genesee Scientific), kým bunky CRFK nedosahovali približne 75-85% konfluenciu. Sériové 10-násobné riedenia sa pripravili z FIPV zásobného roztoku a 200 μl vzoriek z každého riedenia sa pridalo do 10-jamkových replikátov. 72 hodín po infekcii sa bunky fixovali metanolom a zafarbili sa kryštálovou violeťou (Sigma-Aldrich). Jednotlivé jamky sa vizuálne hodnotili na vírusom indukovaný CPE, hodnotili sa ako CPE pozitívne alebo negatívne a TCID50 sa stanovila na základe rovnice log10TCID50 = [celkový počet # jamiek CPE pozitívny / # replikátov] + 0.5, aby reflektovali infekčné virióny na mililiter supernatantu [68].
Kvantifikácia FIPV pomocou qRT-PCR
Bezbunková vírusová RNA bola izolovaná z východiskového vírusu pomocou QIAamp Viral RNA Mini Kit (Qiagen) podľa pokynov výrobcu. Izolovaná RNA bola ošetrená DNázou (Turbo DNáza, Invitrogen) a následne reverzne transkribovaná pomocou súpravy High-Capacity RNA-to-CDNA (Applied Biosystems) podľa protokolov výrobcov. Počet kópií FIPV a mačacej GAPDH cDNA sa stanovil s použitím Real-Time PCR systému Applied Biosystems ‚QuantStudio 3 a PowerUp SYBR Green Master Mix podľa protokolu výrobcu pre 10 μL reakciu. Každá PCR reakcia sa uskutočňovala trikrát s vodným templátom ako negatívnou kontrolou a plazmidovou DNA ako pozitívnou kontrolou. Kontrolná reakcia vylučujúca reverznú transkriptázu bola zahrnutá do každej sady testov PCR v reálnom čase. Templáty cDNA sa amplifikovali pomocou FIPV forward primeru, 5’-GGAAGTTTAGATTTGATTTGGCAATGCTAG a FIP reverzného primeru, 5’-AACAATCACTAGATCCAGACGTTAGCT (terminálna časť génu FIPV 7b) [25]. PCR v reálnom čase pre mačací domáci gén GAPDH sa uskutočňovala súčasne s použitím primérov, 5 GAPDH, 5’-AAATTCCACGGCACAGTCAAG a 3 GAPDH, 5’-TGATGGGCTTTCCATTGATGA. Podmienky cyklovania pre FIPV aj GAPDH amplikóny boli nasledujúce: 50°C počas 2 minút, 95°C počas 2 minút, potom nasledovalo 40 cyklov pri teplote 95°C počas 15s, 58°C počas 30s, 72°C počas 1 minúty. Posledný krok obsahoval disociačnú krivku na vyhodnotenie špecifičnosti väzby priméru. Počet kópií FIPV a GAPDH bol vypočítaný na základe štandardných kriviek generovaných v našom laboratóriu. Kópie cIPNA FIPV stanovené pomocou RT PCR v reálnom čase sa normalizovali na 106 kópií mačacej GAPDH cDNA.
Vývoj anti-helikázových chemických fragmentov
Lieky skúmané a opísané v tejto štúdii boli už známe antivírusové látky. Naproti tomu bol helikázový enzým FIPV klonovaný, exprimovaný a použitý ako cieľ pre koronavírus a enzýmovo špecifickým vírusovým objavom. Cieľová sekvencia DNA AviTag-FIP Helicase-HisTag bola optimalizovaná a syntetizovaná. Syntetizovaná sekvencia bola klonovaná (Adeyemi Adedeji) do vektora pET30a s tagom Avi-His na expresiu proteínu v E. coli. Kmeň E. coli BL21 (DE3) sa transformoval rekombinantným plazmidom. Jedna kolónia sa naočkovala do 1 litra auto-indukovaného média obsahujúceho antibiotikum a kultúra sa inkubovala pri 37°C pri 200 ot./min.
Keď OD600 dosiahol asi 3, teplota bunkovej kultúry bola na 16 hodín zmenená na 15°C. Bunky sa zozbierali centrifugáciou. Bunkové pelety sa resuspendovali v lyzačnom pufri s následnou sonifikáciou. Zrazenina po centrifugácii sa rozpustila pomocou denaturačného činidla. Cieľový proteín sa získal jednostupňovým čistením na Ni kolóne. Cieľový proteín sa sterilizoval 0,22 um filtrom. Výťažok bol 7,2 mg pri 0,90 mg/ml a bol uložený v PBS, 10% glycerole, 0,5 mM L-arginíne, pH 7,4. Koncentrácia bola stanovená Bradfordovým proteínovým testom s BSA ako štandardom. Čistota proteínu a molekulová hmotnosť sa stanovili pomocou SDS-PAGE s potvrdením Western blot.
Skríning fragmentov povrchovej plazmónovej rezonancie (SPR) sa uskutočňoval na platforme ForteBio Pioneer FE SPR. Použil sa senzorový čip HisCap, ktorý obsahuje povrchovú matricu NTA. Kanály 1 a 3 boli naplnené 100uM NiCI2, nasledovalo injekčné podanie 50ug/ml FIP proteínu. Kanál 2 bol ponechaný bez bielkovín, rovnako ako NiCl2, ako referencia. Kanál 1 bol imobilizovaný na hustotu ~8000 RU, zatiaľ čo kanál 3 obsahoval približne 12000 RU. Bol použitý kanál 1. Pufrom použitým na imobilizáciu bol 10 mM HEPES, pH 7.4, 150mM NaCI a 0,1% Tween-20. Pre tento test sa pridal DMSO do konečnej koncentrácie 4%. Proprietárna knižnica zlúčenín sa zriedila v rovnakom tlmivom roztoku bez DMSO na konečnú koncentráciu DMSO 4% DMSO. Zlúčeniny z knižnice sa testovali pri koncentrácii 100 uM s použitím gradientovej injekčnej metódy OneStep. Nálezy boli vybrané na základe RU a kinetiky a použité pre bunkový skríning.
Vírusový plaque (doštičkový) test
Na skríning antivírusovej aktivity zlúčenín boli infikované bunky CRFK ošetrené zlúčeninami v šiestich jamkových replikátoch a porovnávané s jamkami pozitívnej kontroly (infikované bunky), negatívnymi kontrolami (neinfikované bunky) a kontrolami liečby (infikované bunky ošetrené známou účinnou antivírusovou zlúčeninou) súčasne na každej platni pre tkanivové kultúry. CRFK bunky sa pestovali na 96-jamkových platniach pre tkanivové kultúry (Genesee Scientific) obsahujúcich 200 ul kultivačného média. Pri ~75-85% konfluencii buniek bolo médium v neinfikovaných kontrolných jamkách odsaté a nahradené 200 ul čerstvého média. Médium v infikovaných jamkách bolo odsaté a nahradené médiom inokulovaným FIPV pri multiplicite infekcie (MOI) 0,004 infekčného viriónu na bunku. Doštička pre tkanivovú kultúru sa inkubovala 1 hodinu s periodickým jemným miešaním (manipulácie typu „číslo osem“) uskutočňovaným každých 15 minút, aby sa uľahčila interakcia vírus-bunka. Po 1 hodine po infekcii sa každá predpokladaná antivírusová zlúčenina pridala do šiestich jamiek infikovaných FIPV (na stanovenie antivírusovej účinnosti zlúčeniny) a šiestich neinfikovaných kontrolných jamiek (na skríning cytotoxicity zlúčeniny v bunkách CRFK). Všetky zlúčeniny boli pôvodne skrínované pri 10 μM, s výnimkou zlúčenín „chemického fragmentu“ dodávaných M. Olsenom (Midwestern University), ktoré boli hodnotené pri 50 μM. Doštičky s tkanivovými kultúrami sa inkubovali 72 hodín pri 37°C a potom sa zafixovali metanolom a zafarbili sa kryštálovou fialovou farbou. Doštičky sa skenovali na absorbanciu pri 620 nm pomocou čítačky doštičiek ELISA (FilterMax F3, Molecular Devices; Softmax Pro, Molecular Devices). Pre každú liečebnú podmienku sa zaznamenali jednotlivé hodnoty absorbancie jamky spolu s priemernou hodnotou absorbancie a strednou chybou priemeru pre 6 jamkové experimentálne replikáty.
Pre látky, ktoré preukázali antivírusovú účinnosť pri počiatočnom skríningu pri 10 alebo 50μM (chránené pred vírusom asociovaným CPE), sa EC50 stanovila vykonaním série progresívnych 2-násobných riedení zlúčenín vo vírusovom plaque teste. Na stanovenie EC50 sa bunky CRFK pestovali na 96-jamkových doštičkách pre tkanivové kultúry podobne ako v prípade antivírusového skríningového testu. Okrem neinfikovaných kontrolných jamiek boli všetky zostávajúce jamky infikované FIPV, ako je opísané vyššie. Séria dvojnásobného riedenia sa pohybovala od 20μM do 0μM a každá koncentrácia sa uskutočňovala v šiestich jamkových replikátoch. Počet krokov riedenia bol v rozmedzí od 6 do 14 a bol závislé od zlúčeniny. Šesť jamkových replikátov neinfikovaných buniek CRFK slúžilo ako kontrola pre normálne bunky CRFK; šesť replikátov CRFK buniek infikovaných FIPV slúžilo ako neošetrené kontrolné jamky infikované FIPV; a šesť jamkových replikátov FIPV infikovaných CRFK buniek ošetrených GS-441524 slúžilo ako kontrolné jamky na ochranu pred vírusom indukovanou bunkovou smrťou na základe publikovaných údajov týkajúcich sa účinnosti použitia GS-441524 in vitro v CRFK bunkách [26].
Platne s tkanivovými kultúrami sa inkubovali 72 hodín a potom sa zafixovali metanolom, zafarbili sa kryštálovou violeťou a pomocou čítačky doštičiek ELISA sa skenovala absorbancia pri 620 nm. Jednotlivé hodnoty absorbancie spolu s priemernou hodnotou absorbancie a štandardnou odchýlkou pre 6 jamkové experimentálne replikáty boli zaznamenané pre každú podmienku liečby. EC50 bola vypočítaná vynesením nelineárnej regresnej rovnice (krivka dávka-odpoveď) pomocou programu Prism 8 (GraphPad).
Vírusový RNA knock-down test
Na kvantifikáciu inhibície produkcie vírusovej RNA zlúčeninou sa použili testy v reálnom čase RT-PCR. CRFK bunky sa kultivovali na 6-jamkovej doštičke pre tkanivové kultúry (Genesee Biotek). Pri približne 75-85% splývaní buniek bolo kultivačné médium nahradené čerstvým médiom a bunky boli infikované FIPV sérotypu II pri MOI 0,2 (MOI na základe TCID50 biotestu/pfu). Doštičky sa inkubovali jednu hodinu s periodickým jemným trepaním každých 15 minút. Jamky infikované FIPV boli ošetrené jednou (monoterapia), dvoma alebo tromi (kombinovaná antikoravírusová terapia) antivírusovými zlúčeninami; každé experimentálne ošetrenie sa uskutočňovalo trikrát. Dávka zlúčeniny bola založená na EC50 zlúčenín a pohybovala sa od 0,001 do 20 μM. Pre každú experimentálnu sadu slúžili tri kultivačné jamky s FIPV-infikovanými a neošetrenými CRFK bunkami ako vírusom infikované kontroly. Infikované bunkové kultúry boli následne inkubované po dobu 24 hodín a celková RNA spojená s bunkami bola izolovaná pomocou mini súpravy PureLink ™ RNA (Invitrogen). RNA bola ošetrená DNAsou (TurboDNAse, Ambion), reverzne transkribovaná na cDNA pomocou súpravy High-Capacity RNA-to-cDNA Kit (Applied Biosystems) a cIPNA FIPV a mačacie GAPDH cDNA boli merané pomocou qRT-PCR v reálnom čase, ako je opísané vyššie . Násobné zníženie vírusového titra sa stanovilo delením normalizovaného priemerného počtu kópií FIPV RNA pre neošetrené bunky CRFK infikované FIPV na normalizovaný priemerný počet kópií FIPV RNA pre liečené bunky CRFK požadovanou zlúčeninou (zlúčeninami). Očakávaný aditívny účinok sa stanovil pridaním násobnej redukcie pre každú monoterapiu použitú v kombinácii. Zložený aditívny účinok sa určil vydelením predikovaného aditívneho účinku na hodnotu kombinovanej redukcie násobku pre konkrétnu kombinovanú liečbu.
Stanovenie bezpečnostných profilov cytotoxicity (CSP).
Cytotoxicita zlúčeniny v mačacích bunkách sa hodnotila pomocou komerčne dostupnej súpravy (CellTox Green Cytotoxicity Assay, Promega) podľa pokynov výrobcu. Neošetrené bunky CRFK sa použili ako negatívne kontroly a bunky sa ošetrili cytotoxickým roztokom poskytnutým výrobcom ako pozitívne kontroly toxicity. Stručne, okrem kontrolných jamiek boli bunky CRFK nanesené na 96-jamkové doštičky pre tkanivové kultúry (Genesee Scientific) v štyroch jamkových replikátoch s 5, 10, 25, 50 alebo 100 μM koncentráciami požadovanej zlúčeniny a boli inkubované po dobu 72 hodín. Po 72 hodinách sa na všetky jamky nanieslo farbivo viažuce sa na súpravu DNA, inkubované pri 37°C chránené pred svetlom po dobu 15 minút a intenzite fluorescencie pri 485-500 nm Ex/520-530 nm EM bola následne stanovená pomocou čítačky doštičiek (FilterMax F3, Molekulárne zariadenia; Softmax Pro, Molekulárne zariadenia). Predpokladalo sa, že cytotoxicita zlúčeniny v konkrétnej koncentrácii je úmerná intenzite fluorescencie na základe selektívnej penetrácie a väzby farbiva na DNA degenerovaných, apoptotických alebo nekrotických buniek. Rozsah cytotoxicity sa stanovil nastavením hodnoty fluorescencie pre bunky ošetrené reagentom pre pozitívnu kontrolu na 100% a neošetrené mačacie bunky ako 0% cytotoxicity. Priemerná hodnota fluorescencie pre štyri jamky obsahujúce každú koncentráciu zlúčeniny sa potom interpolovala ako percento (percento cytotoxicity) v rozmedzí od 0 do 100%.
Konflikt záujmov
Autori prehlasujú, že nedošlo k žiadnemu konfliktu záujmov.
Oceňujeme financovanie poskytované Winn Feline Foundation (MTW 17-020; MTW 19-026) a Kalifornskou univerzitou v Davise, Centre for Companion Animal Health (CCAH; 2018-92-F; 2018-94-FE) prostredníctvom darov určených pre výskum FIP viacerými individuálnymi darcami a organizáciami (SOCK FIP, Davis, CA) a nadáciami (fond Philip Raskin, Kansas City, KS).
Doplnkové informácie
Kompletný zoznam zlúčenín testovaných in vitro na aktivitu proti FIPV
Inhibítory nukleozidovej polymerázy
| 12x GS Nuc Analogs | Nucleoside analog | NPI |
| GS-441524 (China-sourced) | Nucleoside analog Adenosine | NPI |
| 3-Deazaneplanocin A Hydrochloride | Nucleoside analog Adenosine | NPI |
| Adefovir | Nucleoside analog Adenosine | NPI |
| Galidesivir | Nucleoside analog Adenosine | NPI |
| GS-441524 (Manufactured in China) | Nucleoside analog Adenosine | NPI |
| MK-0608 | Nucleoside analog Adenosine | NPI |
| NITD008 | Nucleoside analog Adenosine | NPI |
| Didanosine | Nucleoside analog Adenosine | NPI |
| Tenofovir alafenamide | Nucleoside analog Adenosine | NPI |
| Tenofovir disoproxil fumarate | Nucleoside analog Adenosine | NPI |
| EIDD 1931 | Nucleoside analog Cytidine | |
| EIDD 2801 | Nucleoside analog Cytidine | |
| 2′-C-methylcytidine | Nucleoside analog Cytidine | NPI |
| Gemcitabine Hydrochloride | Nucleoside analog Cytidine | NPI |
| 2-C-methylguanosine | Nucleoside analog Guanosine | NPI |
| 7-methylguanosine | Nucleoside analog Guanosine | NPI |
| Entecavir | Nucleoside analog Guanosine | NPI |
| Mizoribine | Nucleoside analog Guanosine | NPI |
| Ribavirin | Nucleoside analog Guanosine | NPI |
| PSI-6206 | Nucleoside analog Uridine | NPI |
| 6-Azauridine | Nucleoside analog Uridine | NPI |
| Balapiravir | Nucleoside analog Cytidine | NPI |
| Sofosbuvir | Nucleoside analog Uridine | NPI |
| Favipiravir | Nucleoside analog Purine | NPI |
Inhibítory proteázy
| Grazoprevir | NS3/4A protease inhibitor | PI |
| Rupintrivir | Rhinoviral 3CP inhib | PI |
| Lopinavir | Antiretroviral PI | PI |
| Ritonavir | Antiretroviral PI | PI |
| Nelfinavir | Antiretroviral PI | PI |
| Disulfiram (tetraethyliuram disulfide) | Papain-like protease inhib | PI |
| K777/K11777 | Cysteine protease inhibitor | PI |
| Telaprevir | NS3/4A protease inhibitor | PI |
| Camostat mesylate | Serine protease inhibitor | PI |
| Paritaprevir | Serine protease inhibitor | PI |
| GC376 | Coronavirus protease inhibitor | PI |
NS5A inhibítory
| Velpatasvir | NS5A Inhibitor | NS5A Inhibitor |
| Ravidasvir/PPI-668 | NS5A Inhibitor | NS5A Inhibitor |
| Ledipasvir | NS5A Inhibitor | NS5A Inhibitor |
| Ombitasvir | NS5A Inhibitor | NS5A Inhibitor |
| Pibrentasvir | NS5A Inhibitor | NS5A Inhibitor |
| Daclatasvir | NS5A Inhibitor | NS5A Inhibitor |
| Elbasvir | NS5A Inhibitor | NS5A Inhibitor |
NNPI
| Dasabuvir | Non-nucleoside polymerase inhibitor | NNPI |
Iné
| Monensin | Ionophore | Iné |
| Phenazopyridine hydrochloride | Crystalline solid | Iné |
| Pyrvinium pamoate hydrate | Androgen receptor inhibitor | Iné |
| Toremifene citrate | Selective estrogen receptor modulator | Iné |
| AM580 | Retinobenzoic derivative | Iné |
| Homoharringtonine | Translation elongation inhib | Iné |
| Amodiaquine | 4-aminoquinolone | Iné |
Midwestern Chemical Fragments
| F0472-0017 | Midwestern |
| F6190-0257 | Midwestern |
| F6279-0675 | Midwestern |
| F2167-1080 | Midwestern |
| F6190-0740 | Midwestern |
| F6438-2155 | Midwestern |
| F3411-5663 | Midwestern |
| F6233-0011 | Midwestern |
| F9995-2543 | Midwestern |
| F2124-0890 | Midwestern |
| F2711-2577 | Midwestern |
| F2130-0055 | Midwestern |
| F2493-3358 | Midwestern |
| F2459-0974 | Midwestern |
| F2124-0465 | Midwestern |
| F1899-2269 | Midwestern |
| F2185-1982 | Midwestern |
| F2189-0717 | Midwestern |
| F2147-0158 | Midwestern |
| F1371-0192 | Midwestern |
| F2156-0057 | Midwestern |
| F9995-2431 | Midwestern |
| F3349-0218 | Midwestern |
| F2156-0059 | Midwestern |
| F2156-0070 | Midwestern |
| F5856-0194 | Midwestern |
| F2147-0975 | Midwestern |
Literatúra
- Chang HW, de Groot RJ, Egberink HF, et al. Feline infectious peritonitis: Insights into feline coronavirus pathobiogenesis and epidemiology based on genetic analysis of the viral 3c gene. J Gen Virol 2010; 91: 415–420.CrossRefPubMedWeb of ScienceGoogle Scholar
- Pedersen NC, Liu H, Dodd KA, et al. Significance of coronavirus mutants in feces and diseased tissues of cats suffering from feline infectious peritonitis. Viruses 2009; 1: 166–184.CrossRefPubMedWeb of ScienceGoogle Scholar
- Vennema H, Poland A, Foley J, et al. Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology 1998; 243: 150–157.CrossRefPubMedWeb of ScienceGoogle Scholar
- Rottier PJM, Nakamura K, Schellen P, et al. Acquisition of Macrophage Tropism during the Pathogenesis of Feline Infectious Peritonitis Is Determined by Mutations in the Feline Coronavirus Spike Protein. J Virol 2005; 79: 14122–14130.Abstract/FREE Full TextGoogle Scholar
- Diaz JV., Poma R. Diagnosis and clinical signs of feline infectious peritonitis in the central nervous system. Can Vet J 2009; 50: 1091–1093.PubMedGoogle Scholar
- Foley JE, Lapointe JM, Koblik P, et al. Diagnostic features of clinical neurologic feline infectious peritonitis. J Vet Intern Med 1998; 12: 415–423.CrossRefPubMedGoogle Scholar
- Pedersen NC. An update on feline infectious peritonitis: Diagnostics and therapeutics. Vet J 2014; 201: 133–141.CrossRefPubMedGoogle Scholar
- Stiles J. Ocular manifestations of feline viral diseases. Vet J 2014; 201: 166–173.CrossRefPubMedGoogle Scholar
- Pedersen NC. A review of feline infectious peritonitis virus infection: 1963-2008. J Feline Med Surg 2009; 11: 225–258.CrossRefPubMedGoogle Scholar
- Hohdatsu T, Yamada M, Tominaga R, et al. Antibody-Dependent Enhancement of Feline Infectious Peritonitis Virus Infection in Feline Alveolar Macrophages and Human Monocyte Cell Line U937 by Serum of Cats Experimentally or Naturally Infected with Feline Coronavirus. J Vet Med Sci 1998; 60: 49–55.CrossRefPubMedGoogle Scholar
- Takano T, Yamada S, Doki T, et al. Pathogenesis of oral type I feline infectious peritonitis virus (FIPV) infection: Antibodydependent enhancement infection of cats with type I FIPV via the oral route. J Vet Med Sci 2019; 81: 911–915.CrossRefPubMedGoogle Scholar
- Vennema H, de Groot RJ, Harbour DA, et al. Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization. J Virol 1990; 64: 1407–1409.Abstract/FREE Full TextGoogle Scholar
- Fenner’s Veterinary Virology. 2017. Epub ahead of print 2017. DOI: 10.1016/c2013-0-06921-6.CrossRefGoogle Scholar
- Wei X, Wang X, Peng W, et al. The Lancet Respiratory Medicine Clinical characteristics of SARS-CoV-2 infected pneumonia with diarrhea. 4.Google Scholar
- Hosoda T, Sakamoto M, Shimizu H, et al. SARS-CoV-2 enterocolitis with persisting to excrete the virus for about two weeks after recovering from diarrhea: A case report. Infect Control Hosp Epidemiol 2020; 1–4.Google Scholar
- Hamming I, Timens W, Bulthuis MLC, et al. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. 2004; 631–637.Google Scholar
- Zhang J, Wu Y, Wang R, et al. Bioinformatic analysis of reproductive system Bioinformatic analysis reveals that the reproductive system is potentially at risk from SARS-CoV-2. 2020; 1–15.Google Scholar
- Zheng YY, Ma YT, Zhang JY, et al. COVID-19 and the cardiovascular system. Nat Rev Cardiol. Epub ahead of print 2020. DOI: 10.1038/s41569-020-0360-5.CrossRefGoogle Scholar
- Chen L, Deng C, Chen X, et al. Ocular manifestations and clinical characteristics of 534 cases of COVID-19 in China: A cross-sectional study. medRxiv 2020; 2020.03.12.20034678.Google Scholar
- Mao L, Wang M, Chen S, et al. Neurological Manifestations of Hospitalized Patients with COVID-19 in Wuhan, China: a retrospective case series study. medRxiv 2020; 2020.02.22.20026500.Google Scholar
- Chen C, Zhou Y, Wang DW. SARS-CoV-2: a potential novel etiology of fulminant myocarditis. Herz 2020; 10–12.Google Scholar
- Li Y-C, Bai W-Z, Hashikawa T. The neuroinvasive potential of SARS-CoV2 may be at least partially responsible for the respiratory failure of COVID-19 patients. J Med Virol 2020; 24–27.Google Scholar
- Shi J, Wen Z, Zhong G, et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science. Epub ahead of print 2020. DOI: 10.1126/science.abb7015.Abstract/FREE Full TextGoogle Scholar
- Pedersen NC, Kim Y, Liu H, et al. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. J Feline Med Surg 2018; 20: 378–392.CrossRefGoogle Scholar
- Murphy BG, Perron M, Murakami E, et al. The nucleoside analog GS-441524 strongly inhibits feline infectious peritonitis (FIP) virus in tissue culture and experimental cat infection studies. Vet Microbiol 2018; 219: 226–233.Google Scholar
- Pedersen NC, Perron M, Bannasch M, et al. Efficacy and safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis. J Feline Med Surg 2019; 21: 271–281.Google Scholar
- Holshue ML, DeBolt C, Lindquist S, et al. First case of 2019 novel coronavirus in the United States. N Engl J Med 2020; 382: 929–936.CrossRefPubMedGoogle Scholar
- Wang M, Cao R, Zhang L, et al. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res 2020; 30: 269–271.CrossRefPubMedGoogle Scholar
- Nakagawa F, May M, Phillips A. Life expectancy living with HIV: Recent estimates and future implications. Curr Opin Infect Dis 2013; 26: 17–25.CrossRefPubMedGoogle Scholar
- Simonetti FR, Kearney MF. Review: Influence of ART on HIV genetics. Current Opinion in HIV and AIDS. Epub ahead of print 2015. DOI: 10.1097/COH.0000000000000120.CrossRefGoogle Scholar
- Arts EJ, Hazuda DJ. HIV-1 antiretroviral drug therapy. Cold Spring Harb Perspect Med; 2. Epub ahead of print 2012. DOI: 10.1101/cshperspect.a007161.Abstract/FREE Full TextGoogle Scholar
- Kim Y, Liu H, Kankanamalage ACG. Reversal of the Progression of Fatal Coronavirus Infection in Cats by a Broad-Spectrum Coronavirus Protease Inhibitor. 2016; 1–18.Google Scholar
- Kumar GN, Jayanti VK, Johnson MK, et al. Metabolism and disposition of the HIV-1 protease inhibitor lopinavir (ABT-378) given in combination with ritonavir in rats, dogs, and humans. Pharm Res 2004; 21: 1622–1630.CrossRefPubMedWeb of ScienceGoogle Scholar
- Sulejmani N, Jafri SM, Gordon SC. Pharmacodynamics and pharmacokinetics of elbasvir and grazoprevir in the treatment of hepatitis C. Expert Opin Drug Metab Toxicol 2016; 12: 353–361.Google Scholar
- Combination Antiretroviral Therapy for HIV Infection – American Family Physician, https://www.aafp.org/afp/1998/0601/p2789.html (accessed 6 April 2020).Google Scholar
- Julg B. Atripla™ – HIV therapy in one pill. 2008; 4: 573–577.Google Scholar
- Clay PG, Taylor TAH, Glaros AG, et al. “One pill, once daily”: what clinicians need to know about Atripla™. 2008; 4: 291–302.Google Scholar
- Hill A, Van Der Lugt J, Sawyer W, et al. How much ritonavir is needed to boost protease inhibitors? Systematic review of 17 dose-ranging pharmacokinetic trials. Aids 2009; 23: 2237–2245.CrossRefPubMedWeb of ScienceGoogle Scholar
- A. H, G.R. G, R.J. B. Ritonavir: Clinical pharmacokinetics and interactions with other anti-HIV agents. Clin Pharmacokinet 1998; 35: 275–291.CrossRefPubMedWeb of ScienceGoogle Scholar
- Kwo P, Gane EJ, Peng C, et al. Effectiveness of Elbasvir and Grazoprevir Combination, With or Without Ribavirin, for Treatment-Experienced Patients With Chronic Hepatitis C Infection. Gastroenterology 2017; 152: 164–175.e4.Google Scholar
- Zhou Y, Vedantham P, Lu K, et al. Protease inhibitors targeting coronavirus and filovirus entry. Antiviral Res 2015; 116: 76–84.CrossRefPubMedGoogle Scholar
- Alessandro SD, Scaccabarozzi D, Signorini L, et al. The Use of Antimalarial Drugs against Viral Infection. 2020; 1–26.Google Scholar
- Rynes RI, Bernstein HN. Ophthalmologic safety profile of antimalarial drugs. In: Lupus. 1993. Epub ahead of print 1993. DOI: 10.1177/0961203393002001051.CrossRefGoogle Scholar
- Briceño E, Reyes S, Sotelo J. Therapy of glioblastoma multiforme improved by the antimutagenic chloroquine. Neurosurg Focus. Epub ahead of print 2003. DOI: 10.3171/foc.2003.14.2.4.CrossRefPubMedGoogle Scholar
- Ekong M, Igiri A, Ekanem T, et al. The effect of amodiaquine on some brain macromolecules of Wistar rats. Int J Biol Chem Sci. Epub ahead of print 2009. DOI: 10.4314/ijbcs.v2i4.39762.CrossRefGoogle Scholar
- Mackenzie AH. Pharmacologic actions of 4-aminoquinoline compounds. Am J Med. Epub ahead of print 1983. DOI: 10.1016/0002-9343(83)91264-0.CrossRefPubMedWeb of ScienceGoogle Scholar
- Kuroda K. Detection and distribution of chloroquine metabolites in human tissues. J Pharmacol Exp Ther.Google Scholar
- Maguire A, Kolb H. THE EFFECT OF A SYNTHETIC ANTIMALARIAL (AMODIAQUINE) ON THE RETINA. Br J Dermatol. Epub ahead of print 1964. DOI: 10.1111/j.1365-2133.1964.tb15487.x.CrossRefPubMedWeb of ScienceGoogle Scholar
- McAnally D, Siddiquee K, Gomaa A, et al. Repurposing antimalarial aminoquinolines and related compounds for treatment of retinal neovascularization. PLoS One. Epub ahead of print 2018. DOI: 10.1371/journal.pone.0202436.CrossRefGoogle Scholar
- Keyaerts E, Li S, Vijgen L, et al. Antiviral activity of chloroquine against human coronavirus OC43 infection in newborn mice. Antimicrob Agents Chemother 2009; 53: 3416–3421.Abstract/FREE Full TextGoogle Scholar
- Savarino A, Boelaert JR, Cassone A, et al. Effects of chloroquine on viral infections: An old drug against today’s diseases? Lancet Infect Dis 2003; 3: 722–727.CrossRefPubMedWeb of ScienceGoogle Scholar
- Azuma P, Massaquoi M, Job D, et al. Effect of Artesunate–Amodiaquine on Mortality Related to Ebola Virus Disease. 2016; 23–32.Google Scholar
- Boonyasuppayakorn S, Reichert ED, Manzano M, et al. HHS Public Access. 2015; 125–134.Google Scholar
- Baba M, Toyama M, Sakakibara N. Establishment of an antiviral assay system and identification of severe fever with thrombocytopenia syndrome virus inhibitors. 2017; 25: 83–89.Google Scholar
- Savarino A, Lucia MB, Rastrelli E, et al. Anti-HIV Effects of Chloroquine. JAIDS J Acquir Immune Defic Syndr. Epub ahead of print 2004. DOI: 10.1097/00126334-200403010-00002.CrossRefPubMedWeb of ScienceGoogle Scholar
- Al-bari AA. Targeting endosomal acidification by chloroquine analogs as a promising strategy for the treatment of emerging viral diseases. 2017; 5: 1–13.Google Scholar
- Terauchi M. Pharmacokinetics of selective estrogen receptor modulators(SERMs). Clin Calcium 2016; 26: 1571–1581.Google Scholar
- Agents A, Dyall J, Diseases I, et al. Repurposing of Clinically Developed Drugs for Treatment of Middle. Epub ahead of print 2014. DOI: 10.1128/AAC.03036-14.Abstract/FREE Full TextGoogle Scholar
- Zhao Y, Ren J, Harlos K, et al. virus glycoprotein. Nature 2016; 535: 169–172.CrossRefPubMedGoogle Scholar
- Dyall J, Nelson EA, Dewald LE, et al. Identification of Combinations of Approved Drugs With Synergistic Activity Against Ebola Virus in Cell Cultures. 2018; 22908: 672–678.Google Scholar
- Michimae Y, Mikami S, Okimoto K, et al. The First Case of Feline Infectious Peritonitis-like Pyogranuloma in a Ferret Infected by Coronavirus in Japan. 2010; 99–101.Google Scholar
- Ramis A, Amarilla SP. Coronavirus Infection in Ferrets : Antigen Distribution and Inflammatory Response. 2016; 53: 1180–1186.Google Scholar
- Watanabe R, Eckstrand C, Liu H, et al. Characterization of peritoneal cells from cats with experimentally ‑ induced feline infectious peritonitis (FIP) using RNA ‑ seq. Vet Res 2018; 1–15.Google Scholar
- Inhibitor P, Hoffmann M, Kleine-weber H, et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Article SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. 2020; 1–10.Google Scholar
- Zhang B, Zhou X, Zhu C, et al. Immune phenotyping based on neutrophil-to-lymphocyte ratio and IgG predicts disease severity and outcome for patients with COVID-19. medRxiv. Epub ahead of print 2020. DOI: 10.1101/2020.03.12.20035048.Abstract/FREE Full TextGoogle Scholar
- Zhao J, Yuan Q, Wang H, et al. Antibody Responses to SARS-CoV-2 in Patients of Novel Coronavirus Disease 2019. SSRN Electron J. Epub ahead of print 2020. DOI: 10.2139/ssrn.3546052.CrossRefGoogle Scholar
- Cao X. COVID-19: immunopathology and its implications for therapy. Nat Rev Immunol; 2019. Epub ahead of print 2020. DOI: 10.1038/s41577-020-0308-3.CrossRefPubMedGoogle Scholar
- Ramakrishnan MA. Determination of 50% endpoint titer using a simple formula. World J Virol 2016; 5: 85.CrossRefPubMedGoogle Scholar